

Mechanotransduction is required for establishing and maintaining mature inner hair cells and regulating efferent innervation
- PMID: 30275467
- PMCID: PMC6167318
- DOI: 10.1038/s41467-018-06307-w
Mechanotransduction is required for establishing and maintaining mature inner hair cells and regulating efferent innervation
Abstract
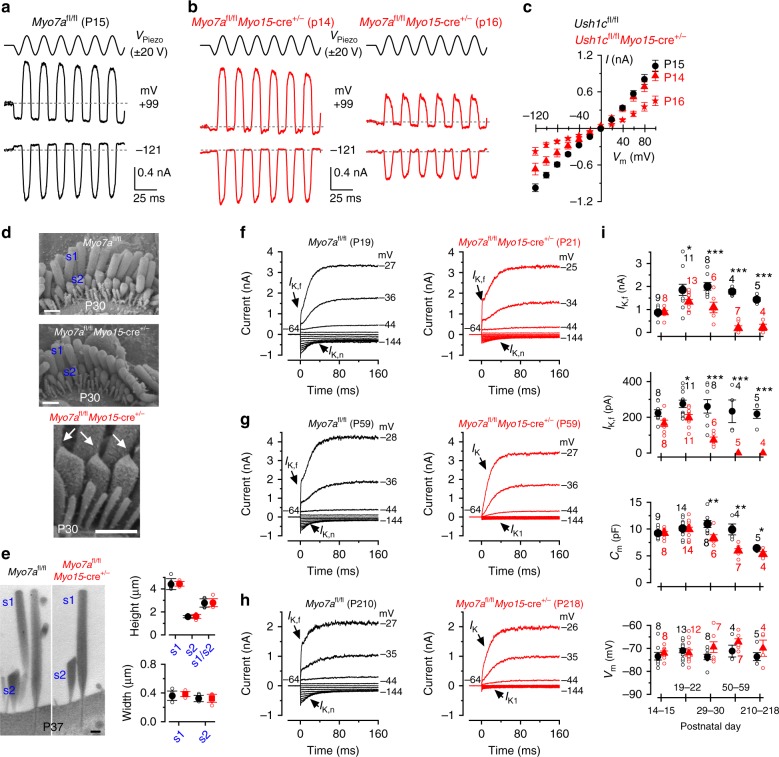
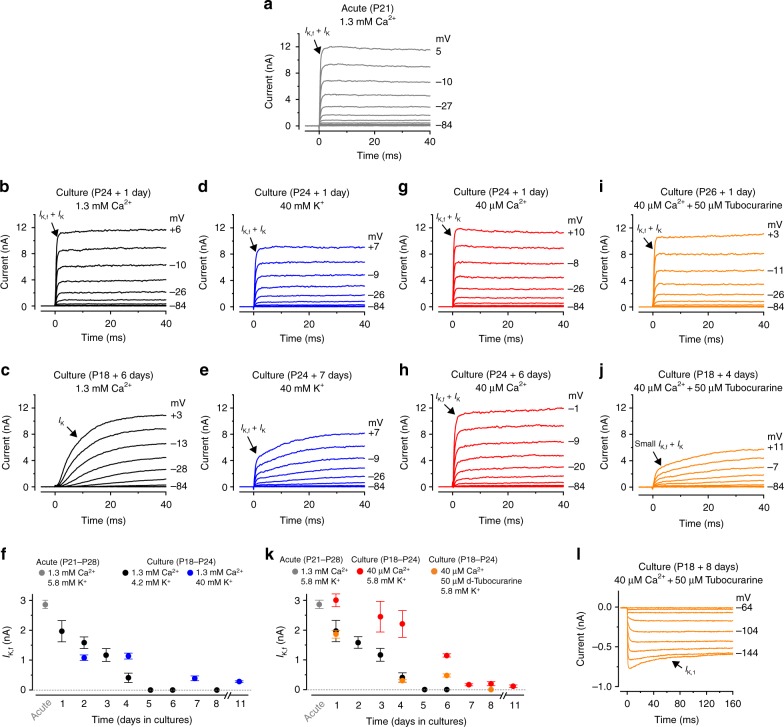
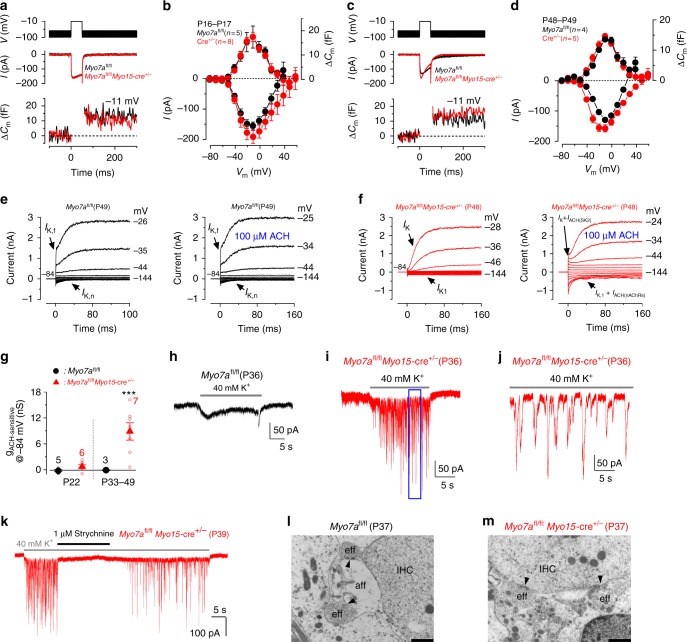
In the adult auditory organ, mechanoelectrical transducer (MET) channels are essential for transducing acoustic stimuli into electrical signals. In the absence of incoming sound, a fraction of the MET channels on top of the sensory hair cells are open, resulting in a sustained depolarizing current. By genetically manipulating the in vivo expression of molecular components of the MET apparatus, we show that during pre-hearing stages the MET current is essential for establishing the electrophysiological properties of mature inner hair cells (IHCs). If the MET current is abolished in adult IHCs, they revert into cells showing electrical and morphological features characteristic of pre-hearing IHCs, including the re-establishment of cholinergic efferent innervation. The MET current is thus critical for the maintenance of the functional properties of adult IHCs, implying a degree of plasticity in the mature auditory system in response to the absence of normal transduction of acoustic signals.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
