

Targeting nuclear receptor NR4A1-dependent adipocyte progenitor quiescence promotes metabolic adaptation to obesity
- PMID: 30277475
- PMCID: PMC6205397
- DOI: 10.1172/JCI98353
Targeting nuclear receptor NR4A1-dependent adipocyte progenitor quiescence promotes metabolic adaptation to obesity
Abstract
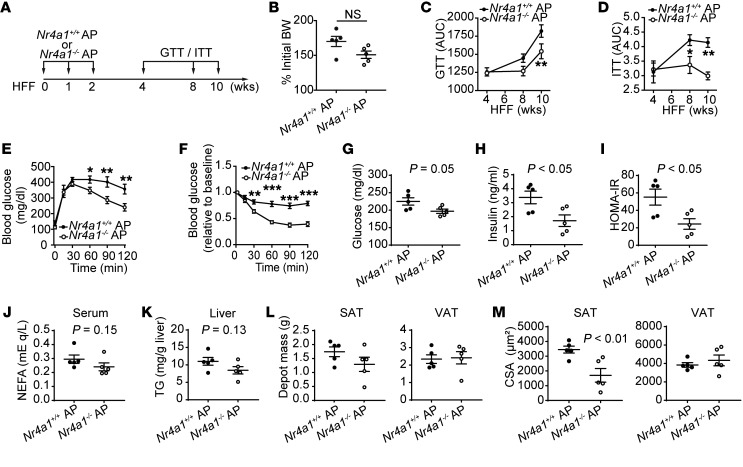
Adipocyte turnover in adulthood is low, suggesting that the cellular source of new adipocytes, the adipocyte progenitor (AP), resides in a state of relative quiescence. Yet the core transcriptional regulatory circuitry (CRC) responsible for establishing a quiescent state and the physiological significance of AP quiescence are incompletely understood. Here, we integrate transcriptomic data with maps of accessible chromatin in primary APs, implicating the orphan nuclear receptor NR4A1 in AP cell-state regulation. NR4A1 gain and loss of function in APs ex vivo decreased and enhanced adipogenesis, respectively. Adipose tissue of Nr4a1-/- mice demonstrated higher proliferative and adipogenic capacity compared with that of WT mice. Transplantation of Nr4a1-/- APs into the subcutaneous adipose tissue of WT obese recipients improved metrics of glucose homeostasis relative to administration of WT APs. Collectively, these data identify NR4A1 as a previously unrecognized constitutive regulator of AP quiescence and suggest that augmentation of adipose tissue plasticity may attenuate negative metabolic sequelae of obesity.
Keywords: Adipose tissue; Cell Biology; Diabetes; Metabolism; Obesity.
Conflict of interest statement
Figures







References
-
- Mokdad AH, et al. Prevalence of obesity, diabetes, and obesity-related health risk factors, 2001. JAMA. 2003;289(1):76–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
