

Functional trait divergence and trait plasticity confer polyploid advantage in heterogeneous environments
- PMID: 30281801
- PMCID: PMC6587808
- DOI: 10.1111/nph.15508
Functional trait divergence and trait plasticity confer polyploid advantage in heterogeneous environments
Abstract
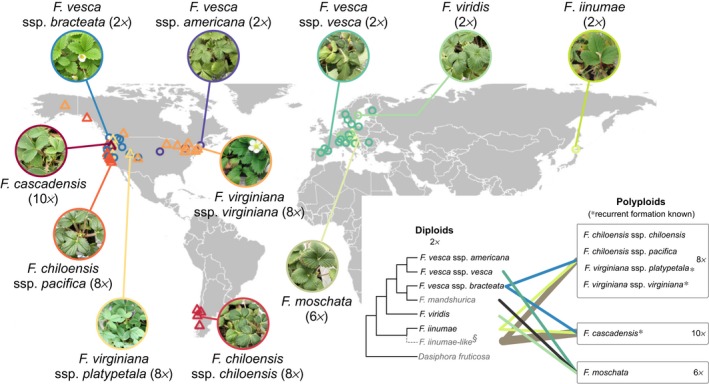
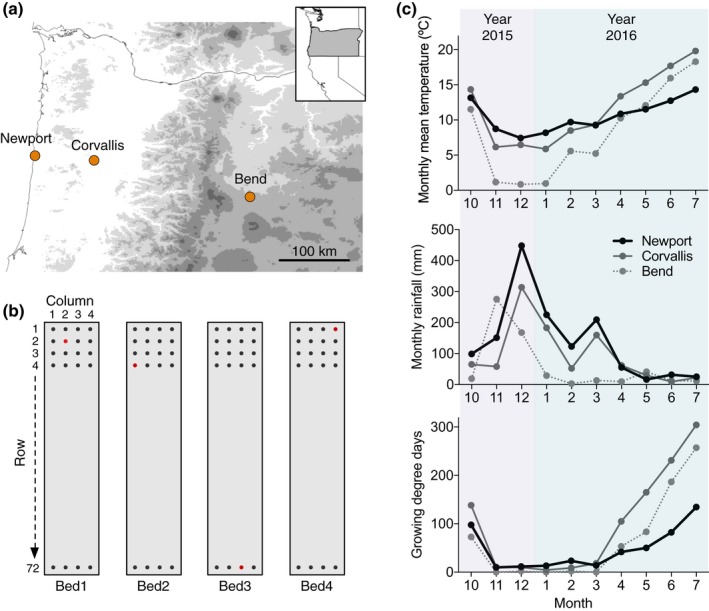
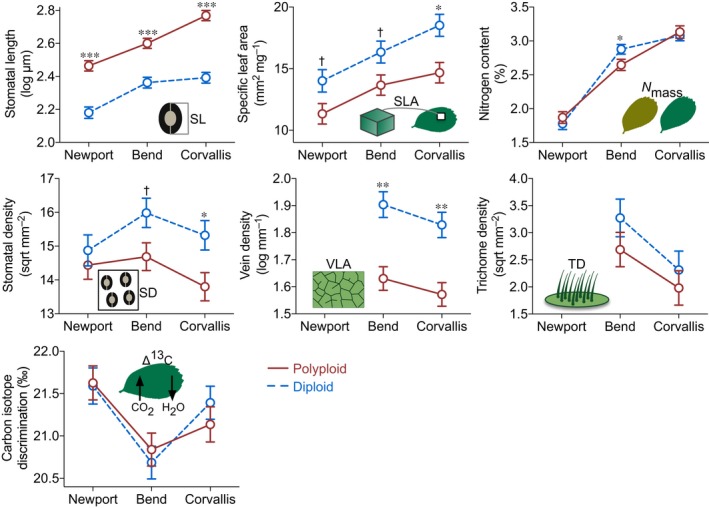
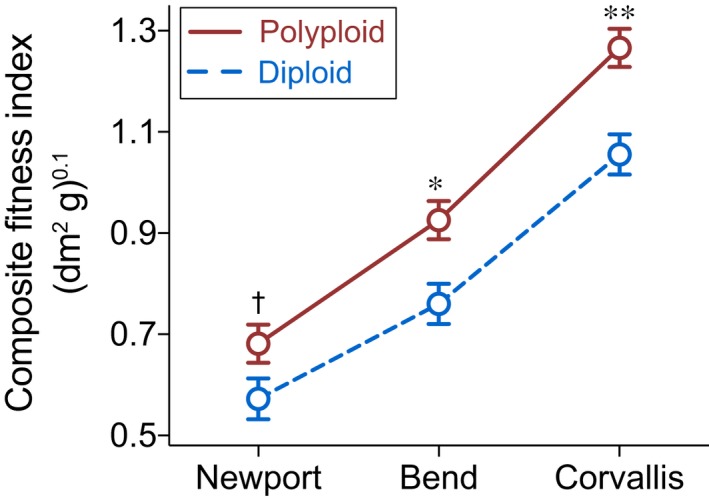
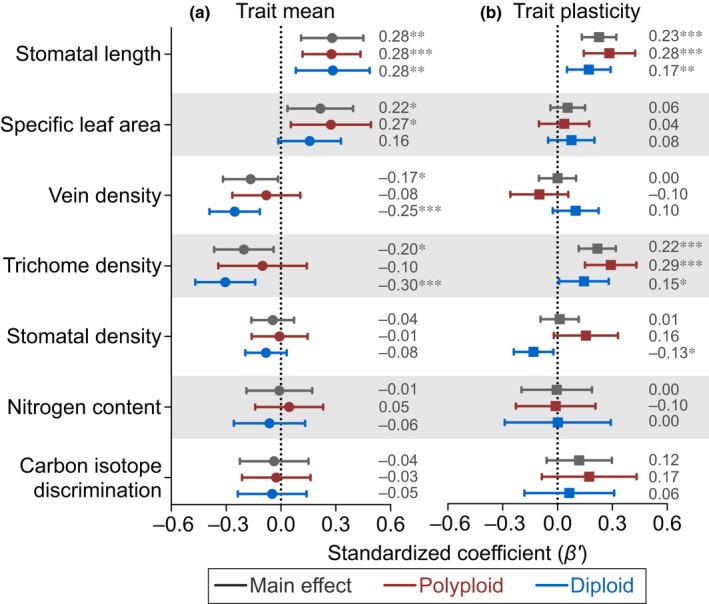
Polyploidy, or whole-genome duplication often with hybridization, is common in eukaryotes and is thought to drive ecological and evolutionary success, especially in plants. The mechanisms of polyploid success in ecologically relevant contexts, however, remain largely unknown. We conducted an extensive test of functional trait divergence and plasticity in conferring polyploid fitness advantage in heterogeneous environments, by growing clonal replicates of a worldwide genotype collection of six allopolyploid and five diploid wild strawberry (Fragaria) taxa in three climatically different common gardens. Among leaf functional traits, we detected divergence in trait means but not plasticities between polyploids and diploids, suggesting that increased genomic redundancy in polyploids does not necessarily translate into greater trait plasticity in response to environmental change. Across the heterogeneous garden environments, however, polyploids exhibited fitness advantage, which was conferred by both trait means and adaptive trait plasticities, supporting a 'jack-and-master' hypothesis for polyploids. Our findings elucidate essential ecological mechanisms underlying polyploid adaptation to heterogeneous environments, and provide an important insight into the prevalence and persistence of polyploid plants.
Keywords: adaptation; adaptive plasticity; common gardens; functional traits; polyploidy; wild strawberry.
No claim to original US government works New Phytologist © 2018 New Phytologist Trust.
Figures
References
-
- Adams KL, Wendel JF. 2005. Novel patterns of gene expression in polyploid plants. Trends in Genetics 21: 539–543. - PubMed
-
- Ashman T‐L. 2005. The limits on sexual dimorphism in vegetative traits in a gynodioecious plant. The American Naturalist 166: S5–S16. - PubMed
-
- Baker HG. 1965. Characteristics and modes of origin of weeds In: Baker HG, Stebbins LG, eds. The genetics of colonizing species. New York, NY, USA: Academic Press, 147–172.
-
- Balao F, Herrera J, Talavera S. 2011. Phenotypic consequences of polyploidy and genome size at the microevolutionary scale: a multivariate morphological approach. New Phytologist 192: 256–265. - PubMed
-
- Bardil A, de Almeida JD, Combes MC, Lashermes P, Bertrand B. 2011. Genomic expression dominance in the natural allopolyploid Coffea arabica is massively affected by growth temperature. New Phytologist 192: 760–774. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
