

Transsynaptic Binding of Orphan Receptor GPR179 to Dystroglycan-Pikachurin Complex Is Essential for the Synaptic Organization of Photoreceptors
- PMID: 30282023
- PMCID: PMC6203450
- DOI: 10.1016/j.celrep.2018.08.068
Transsynaptic Binding of Orphan Receptor GPR179 to Dystroglycan-Pikachurin Complex Is Essential for the Synaptic Organization of Photoreceptors
Abstract
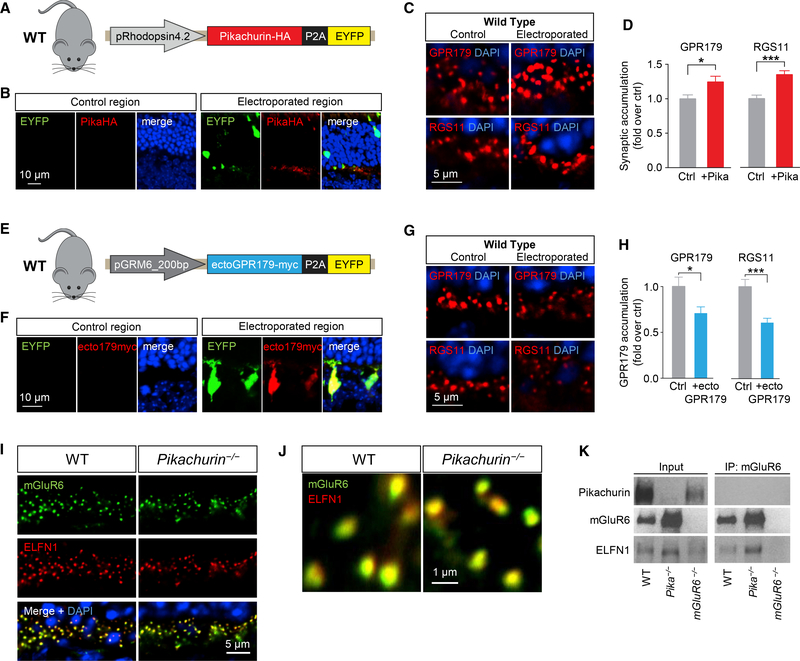
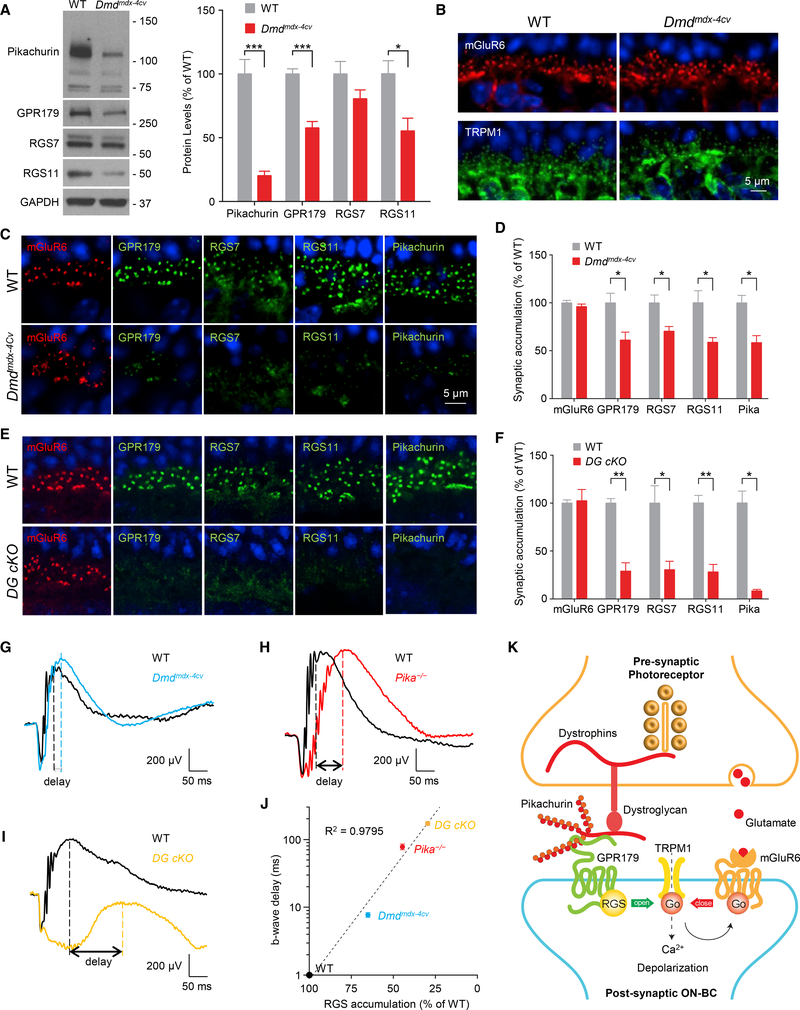
Establishing synaptic contacts between neurons is paramount for nervous system function. This process involves transsynaptic interactions between a host of cell adhesion molecules that act in cooperation with the proteins of the extracellular matrix to specify unique physiological properties of individual synaptic connections. However, understanding of the molecular mechanisms that generate functional diversity in an input-specific fashion is limited. In this study, we identify that major components of the extracellular matrix proteins present in the synaptic cleft-members of the heparan sulfate proteoglycan (HSPG) family-associate with the GPR158/179 group of orphan receptors. Using the mammalian retina as a model system, we demonstrate that the HSPG member Pikachurin, released by photoreceptors, recruits a key post-synaptic signaling complex of downstream ON-bipolar neurons in coordination with the pre-synaptic dystroglycan glycoprotein complex. We further demonstrate that this transsynaptic assembly plays an essential role in synaptic transmission of photoreceptor signals.
Keywords: HSPG; ON-bipolar cells; heparan sulfate proteoglycan; orphan receptor; retina; ribbon synapse; synaptic specificity; synaptic transmission; transsynaptic interactions; vision.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Similar articles
-
Structure of the photoreceptor synaptic assembly of the extracellular matrix protein pikachurin with the orphan receptor GPR179.Sci Signal. 2023 Jul 25;16(795):eadd9539. doi: 10.1126/scisignal.add9539. Epub 2023 Jul 25. Sci Signal. 2023. PMID: 37490546 Free PMC article.
-
Pikachurin, a dystroglycan ligand, is essential for photoreceptor ribbon synapse formation.Nat Neurosci. 2008 Aug;11(8):923-31. doi: 10.1038/nn.2160. Epub 2008 Jul 20. Nat Neurosci. 2008. PMID: 18641643
-
Presynaptic dystroglycan-pikachurin complex regulates the proper synaptic connection between retinal photoreceptor and bipolar cells.J Neurosci. 2012 May 2;32(18):6126-37. doi: 10.1523/JNEUROSCI.0322-12.2012. J Neurosci. 2012. PMID: 22553019 Free PMC article.
-
[Essential role of pikachurin, a novel dystroglycan-binding protein, in bipolar dendrite connection to photoreceptor ribbon synapse in the retina].Nippon Ganka Gakkai Zasshi. 2010 Nov;114(11):955-67. Nippon Ganka Gakkai Zasshi. 2010. PMID: 21141075 Review. Japanese.
-
Dystroglycan-HSPG interactions provide synaptic plasticity and specificity.Glycobiology. 2024 Aug 30;34(10):cwae051. doi: 10.1093/glycob/cwae051. Glycobiology. 2024. PMID: 39223703 Free PMC article. Review.
Cited by
-
ELFN2 is a postsynaptic cell adhesion molecule with essential roles in controlling group III mGluRs in the brain and neuropsychiatric behavior.Mol Psychiatry. 2019 Dec;24(12):1902-1919. doi: 10.1038/s41380-019-0512-3. Epub 2019 Sep 4. Mol Psychiatry. 2019. PMID: 31485013 Free PMC article.
-
Loss of ON-Pathway Function in Mice Lacking Lrit3 Decreases Recovery From Lens-Induced Myopia.Invest Ophthalmol Vis Sci. 2024 Sep 3;65(11):18. doi: 10.1167/iovs.65.11.18. Invest Ophthalmol Vis Sci. 2024. PMID: 39250117 Free PMC article.
-
Retinal TRP channels: Cell-type-specific regulators of retinal homeostasis and multimodal integration.Prog Retin Eye Res. 2023 Jan;92:101114. doi: 10.1016/j.preteyeres.2022.101114. Epub 2022 Sep 24. Prog Retin Eye Res. 2023. PMID: 36163161 Free PMC article. Review.
-
Homeostatic Plasticity Shapes the Retinal Response to Photoreceptor Degeneration.Curr Biol. 2020 May 18;30(10):1916-1926.e3. doi: 10.1016/j.cub.2020.03.033. Epub 2020 Apr 2. Curr Biol. 2020. PMID: 32243858 Free PMC article.
-
An Input-Specific Orphan Receptor GPR158-HSPG Interaction Organizes Hippocampal Mossy Fiber-CA3 Synapses.Neuron. 2018 Oct 10;100(1):201-215.e9. doi: 10.1016/j.neuron.2018.08.038. Epub 2018 Oct 2. Neuron. 2018. PMID: 30290982 Free PMC article.
References
-
- Audo I, Bujakowska K, Orhan E, Poloschek CM, Defoort-Dhellemmes S, Drumare I, Kohl S, Luu TD, Lecompte O, Zrenner E, et al. (2012). Whole-exome sequencing identifies mutations in GPR179 leading to autosomal-recessive complete congenital stationary night blindness. Am. J.Hum. Genet 90, 321–330. - PMC - PubMed
-
- Baeg GH, Lin X, Khare N, Baumgartner S, and Perrimon N (2001). Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless. Development 128, 87–94. - PubMed
-
- Bargmann CI, and Marder E (2013). From the connectome to brain function.Nat. Methods 10, 483–490. - PubMed
-
- Bishop JR, Schuksz M, and Esko JD (2007). Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 446, 1030–1037. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
