

Delayed trait development and the convergent evolution of shell kinesis in turtles
- PMID: 30282655
- PMCID: PMC6191694
- DOI: 10.1098/rspb.2018.1585
Delayed trait development and the convergent evolution of shell kinesis in turtles
Abstract
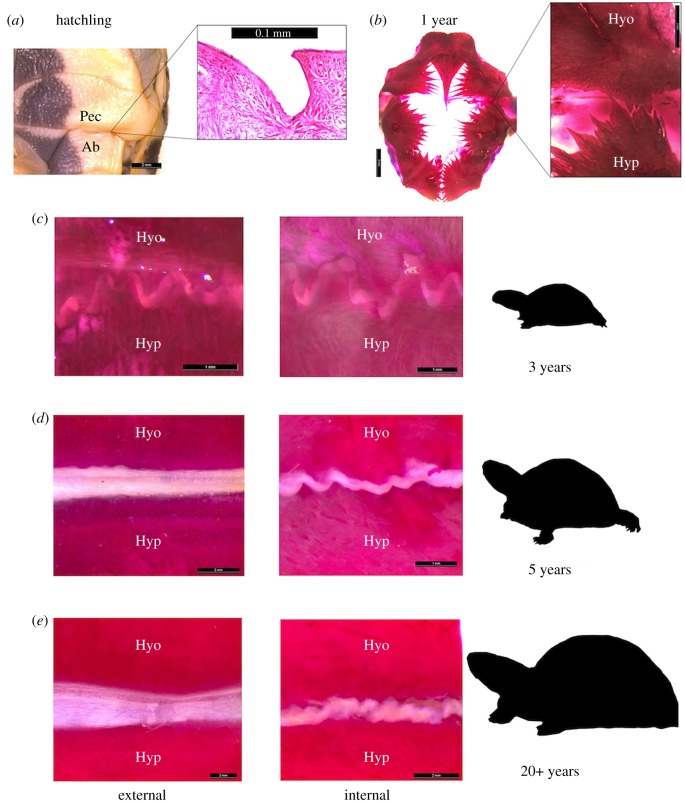
Understanding developmental processes is foundational to clarifying the mechanisms by which convergent evolution occurs. Here, we show how a key convergently evolving trait is slowly 'acquired' in growing turtles. Many functionally relevant traits emerge late in turtle ontogeny, owing to design constraints imposed by the shell. We investigated this trend by examining derived patterns of shell formation associated with the multiple (at least 8) origins of shell kinesis in small-bodied turtles. Using box turtles as a model, we demonstrate that the flexible hinge joint required for shell kinesis differentiates gradually and via extensive repatterning of shell tissue. Disproportionate changes in shell shape and size substantiate that this transformation is a delayed ontogenetic response (3-5 years post-hatching) to structural alterations that arise in embryogenesis. These findings exemplify that the translation of genotype to phenotype may reach far beyond embryonic life stages. Thus, the temporal scope for developmental origins of adaptive morphological change might be broader than generally understood. We propose that delayed trait differentiation via tissue repatterning might facilitate phenotypic diversification and innovation that otherwise would not arise due to developmental constraints.
Keywords: convergent evolution; joint development; ontogeny and phylogeny; shell kinesis; turtle shell evolution.
© 2018 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures



Similar articles
-
Turtle Shell Kinesis Underscores Constraints and Opportunities in the Evolution of the Vertebrate Musculoskeletal System.Integr Org Biol. 2023 Sep 4;5(1):obad033. doi: 10.1093/iob/obad033. eCollection 2023. Integr Org Biol. 2023. PMID: 37840690 Free PMC article. Review.
-
The postembryonic transformation of the shell in emydine box turtles.Evol Dev. 2019 Nov;21(6):297-310. doi: 10.1111/ede.12307. Epub 2019 Aug 23. Evol Dev. 2019. PMID: 31441599
-
Skeletal repatterning enhances the protective capacity of the shell in African hinge-back tortoises (Kinixys).Anat Rec (Hoboken). 2023 Jun;306(6):1558-1573. doi: 10.1002/ar.24954. Epub 2022 May 17. Anat Rec (Hoboken). 2023. PMID: 35582737
-
Skeletal remodelling suggests the turtle's shell is not an evolutionary straitjacket.Biol Lett. 2015 Apr;11(4):20150022. doi: 10.1098/rsbl.2015.0022. Biol Lett. 2015. PMID: 25878046 Free PMC article.
-
Patterning of the turtle shell.Curr Opin Genet Dev. 2017 Aug;45:124-131. doi: 10.1016/j.gde.2017.03.016. Epub 2017 May 29. Curr Opin Genet Dev. 2017. PMID: 28570929 Review.
Cited by
-
Shell biomechanics suggests an aquatic palaeoecology at the dawn of turtle evolution.Sci Rep. 2024 Sep 18;14(1):21822. doi: 10.1038/s41598-024-72540-7. Sci Rep. 2024. PMID: 39294199 Free PMC article.
-
PG-18: turtles reach adult shell shapes at about 65% maximum carapace length.Swiss J Palaeontol. 2025;144(1):47. doi: 10.1186/s13358-025-00395-0. Epub 2025 Aug 5. Swiss J Palaeontol. 2025. PMID: 40778006 Free PMC article.
-
Turtle Shell Kinesis Underscores Constraints and Opportunities in the Evolution of the Vertebrate Musculoskeletal System.Integr Org Biol. 2023 Sep 4;5(1):obad033. doi: 10.1093/iob/obad033. eCollection 2023. Integr Org Biol. 2023. PMID: 37840690 Free PMC article. Review.
References
-
- Dobzhansky T. 1959. Evolution and environment. In The evolution of life: its origin, history, and future (ed. Tax S.), pp. 403–428. Chicago, IL: The University of Chicago Press.
-
- McGhee G. 2011. Convergent evolution: limited forms most beautiful. Cambridge, MA: The MIT Press.
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
