

Into the breach: how cells cope with wounds
- PMID: 30282661
- PMCID: PMC6223217
- DOI: 10.1098/rsob.180135
Into the breach: how cells cope with wounds
Abstract
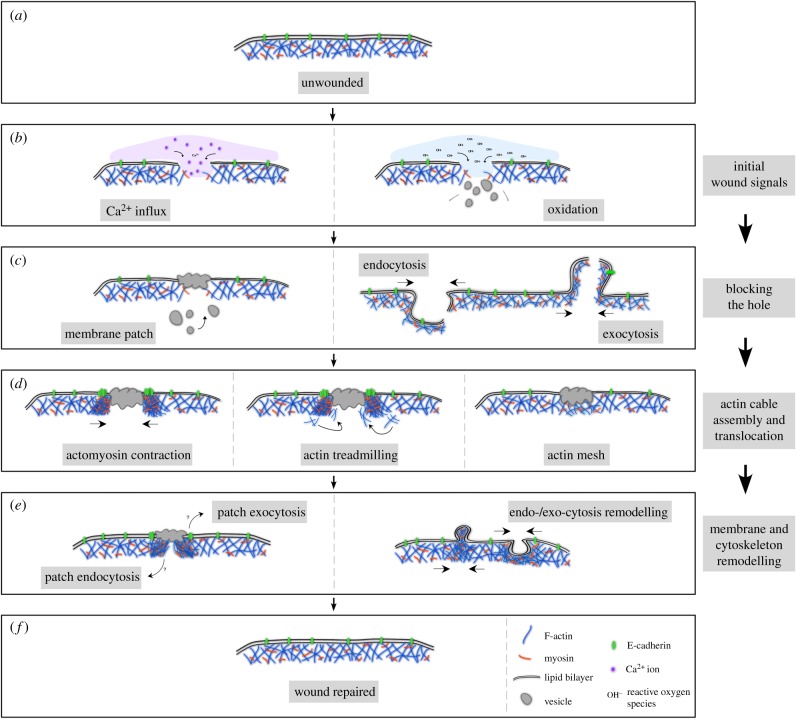
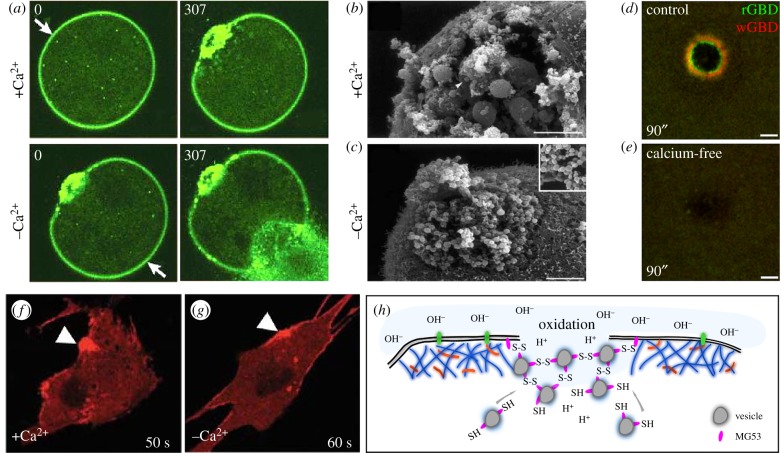
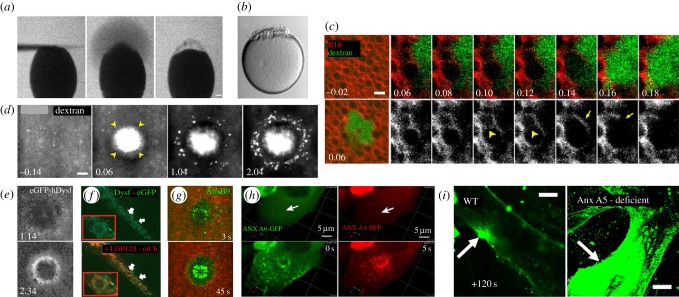
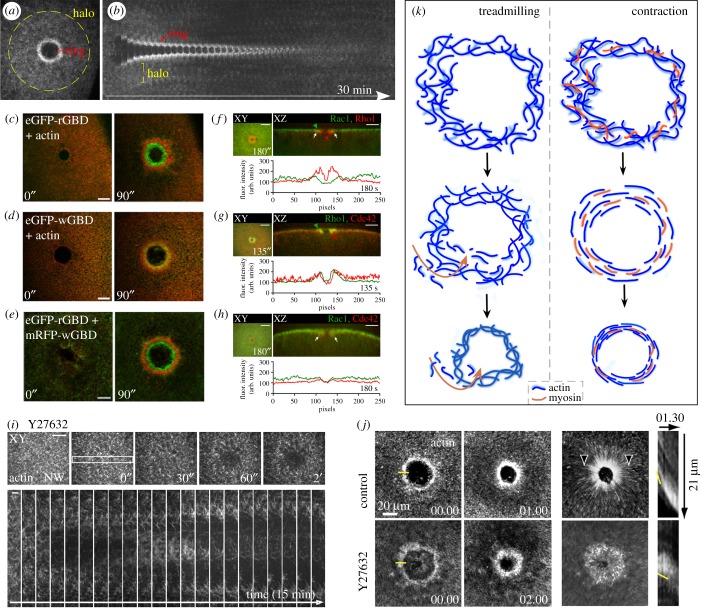
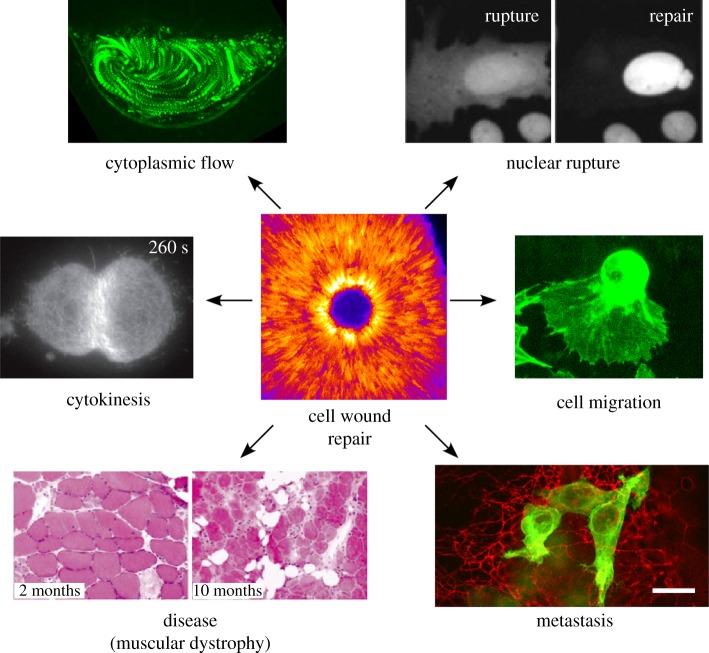
Repair of wounds to individual cells is crucial for organisms to survive daily physiological or environmental stresses, as well as pathogen assaults, which disrupt the plasma membrane. Sensing wounds, resealing membranes, closing wounds and remodelling plasma membrane/cortical cytoskeleton are four major steps that are essential to return cells to their pre-wounded states. This process relies on dynamic changes of the membrane/cytoskeleton that are indispensable for carrying out the repairs within tens of minutes. Studies from different cell wound repair models over the last two decades have revealed that the molecular mechanisms of single cell wound repair are very diverse and dependent on wound type, size, and/or species. Interestingly, different repair models have been shown to use similar proteins to achieve the same end result, albeit sometimes by distinctive mechanisms. Recent studies using cutting edge microscopy and molecular techniques are shedding new light on the molecular mechanisms during cellular wound repair. Here, we describe what is currently known about the mechanisms underlying this repair process. In addition, we discuss how the study of cellular wound repair-a powerful and inducible model-can contribute to our understanding of other fundamental biological processes such as cytokinesis, cell migration, cancer metastasis and human diseases.
Keywords: Rho GTPases; actomyosin; calcium; membrane trafficking; plasma membrane repair; single cell wound repair.
© 2018 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Wrangling Actin Assemblies: Actin Ring Dynamics during Cell Wound Repair.Cells. 2022 Sep 6;11(18):2777. doi: 10.3390/cells11182777. Cells. 2022. PMID: 36139352 Free PMC article. Review.
-
Cytoskeleton responses in wound repair.Cell Mol Life Sci. 2012 Aug;69(15):2469-83. doi: 10.1007/s00018-012-0928-2. Epub 2012 Feb 15. Cell Mol Life Sci. 2012. PMID: 22349211 Free PMC article. Review.
-
Actin Cytoskeletal Dynamics in Single-Cell Wound Repair.Int J Mol Sci. 2021 Oct 8;22(19):10886. doi: 10.3390/ijms221910886. Int J Mol Sci. 2021. PMID: 34639226 Free PMC article. Review.
-
Rehabilitation and the single cell.Curr Opin Cell Biol. 2007 Feb;19(1):95-100. doi: 10.1016/j.ceb.2006.12.001. Epub 2006 Dec 14. Curr Opin Cell Biol. 2007. PMID: 17174083 Free PMC article. Review.
-
Actin dynamics and myosin contractility during plasma membrane repair and restoration: Does one ring really heal them all?Curr Top Membr. 2019;84:17-41. doi: 10.1016/bs.ctm.2019.07.004. Epub 2019 Sep 13. Curr Top Membr. 2019. PMID: 31610862 Review.
Cited by
-
Voltage-gated calcium channels act upstream of adenylyl cyclase Ac78C to promote timely initiation of dendrite regeneration.PLoS Genet. 2024 Aug 26;20(8):e1011388. doi: 10.1371/journal.pgen.1011388. eCollection 2024 Aug. PLoS Genet. 2024. PMID: 39186815 Free PMC article.
-
Autocrine insulin pathway signaling regulates actin dynamics in cell wound repair.PLoS Genet. 2020 Dec 11;16(12):e1009186. doi: 10.1371/journal.pgen.1009186. eCollection 2020 Dec. PLoS Genet. 2020. PMID: 33306674 Free PMC article.
-
Plasma membrane damage limits replicative lifespan in yeast and induces premature senescence in human fibroblasts.Nat Aging. 2024 Mar;4(3):319-335. doi: 10.1038/s43587-024-00575-6. Epub 2024 Feb 22. Nat Aging. 2024. PMID: 38388781 Free PMC article.
-
Wrangling Actin Assemblies: Actin Ring Dynamics during Cell Wound Repair.Cells. 2022 Sep 6;11(18):2777. doi: 10.3390/cells11182777. Cells. 2022. PMID: 36139352 Free PMC article. Review.
-
Creating wounds in cell monolayers using micro-jets.Biomicrofluidics. 2021 Feb 8;15(1):014108. doi: 10.1063/5.0043312. eCollection 2021 Jan. Biomicrofluidics. 2021. PMID: 33598064 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
