

Extracellular Hepatitis B Virus RNAs Are Heterogeneous in Length and Circulate as Capsid-Antibody Complexes in Addition to Virions in Chronic Hepatitis B Patients
- PMID: 30282709
- PMCID: PMC6258948
- DOI: 10.1128/JVI.00798-18
Extracellular Hepatitis B Virus RNAs Are Heterogeneous in Length and Circulate as Capsid-Antibody Complexes in Addition to Virions in Chronic Hepatitis B Patients
Abstract
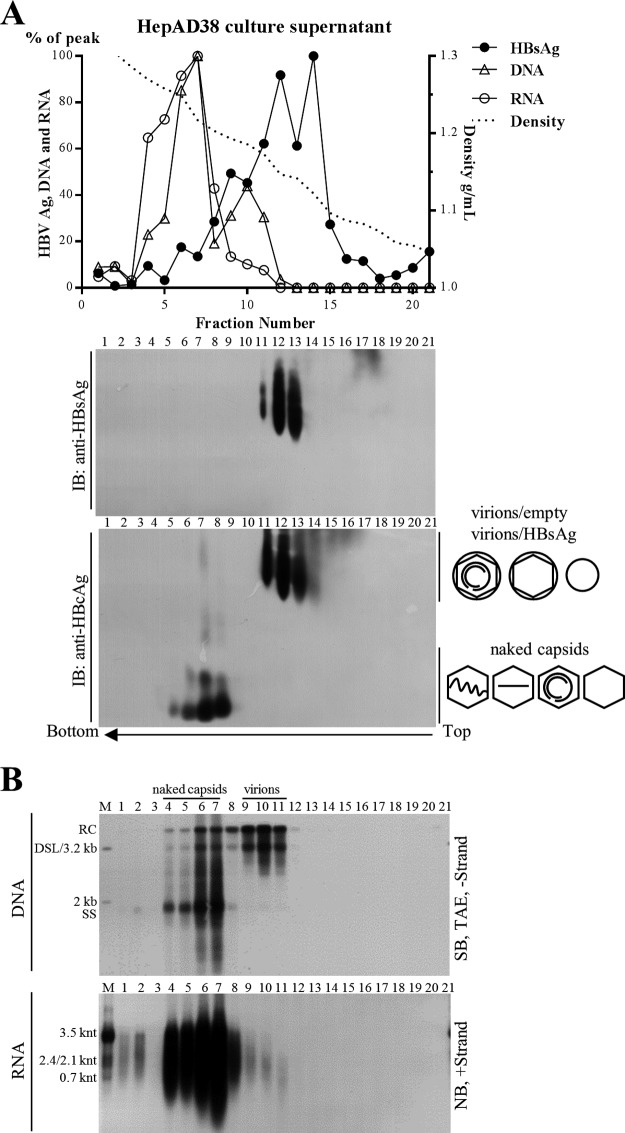
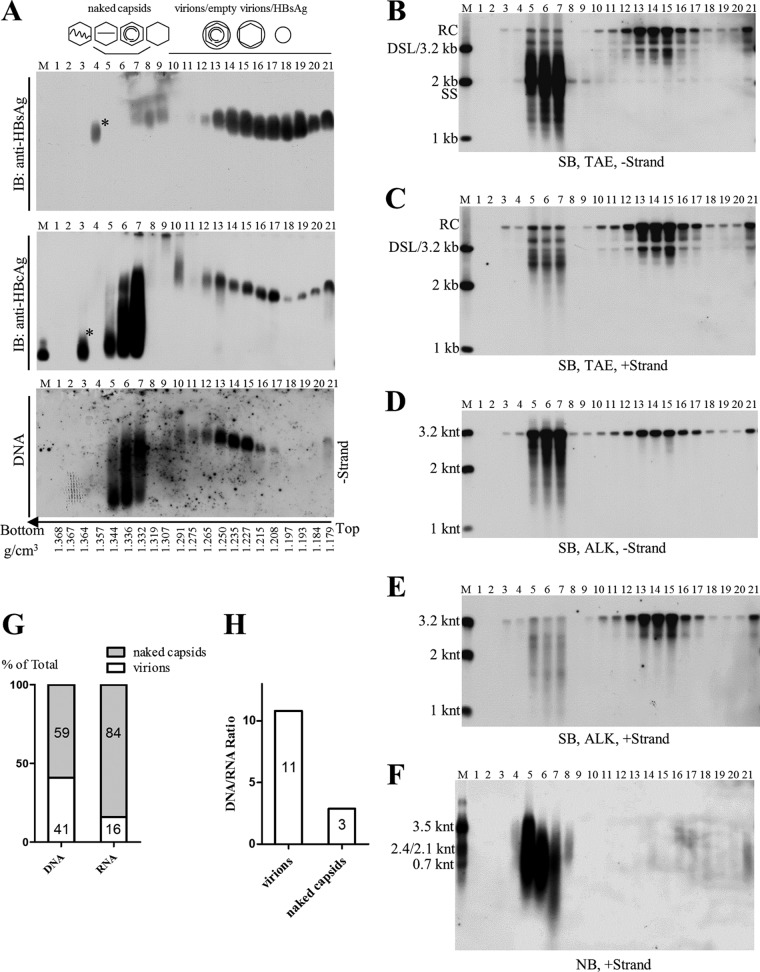
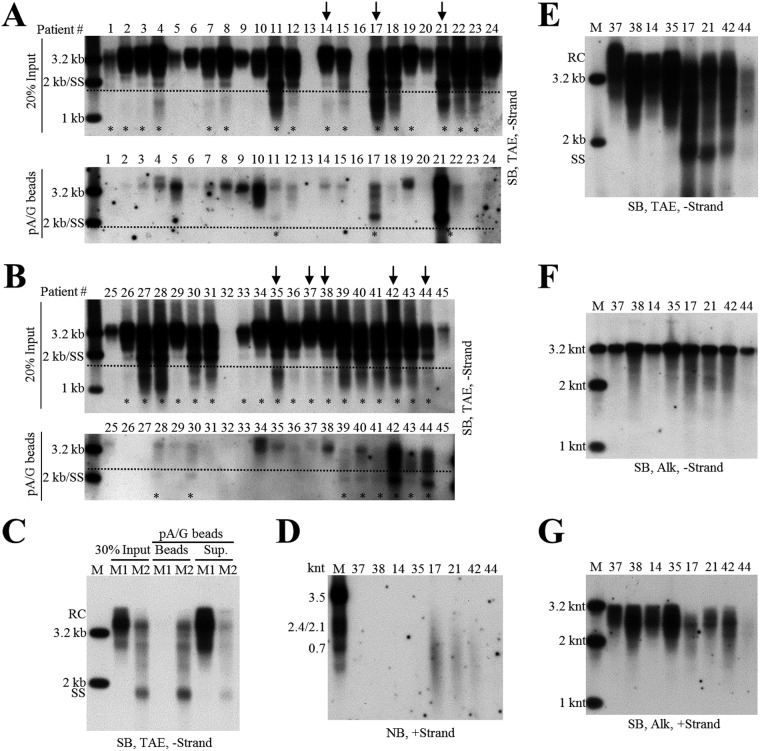
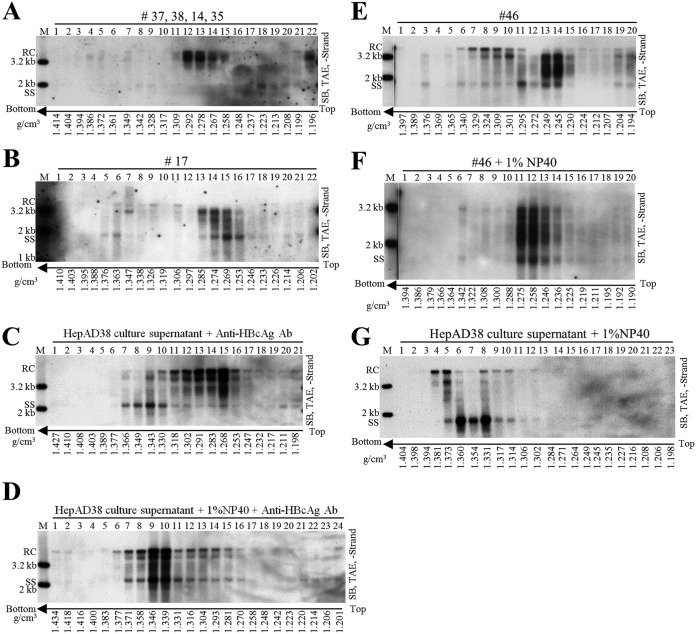
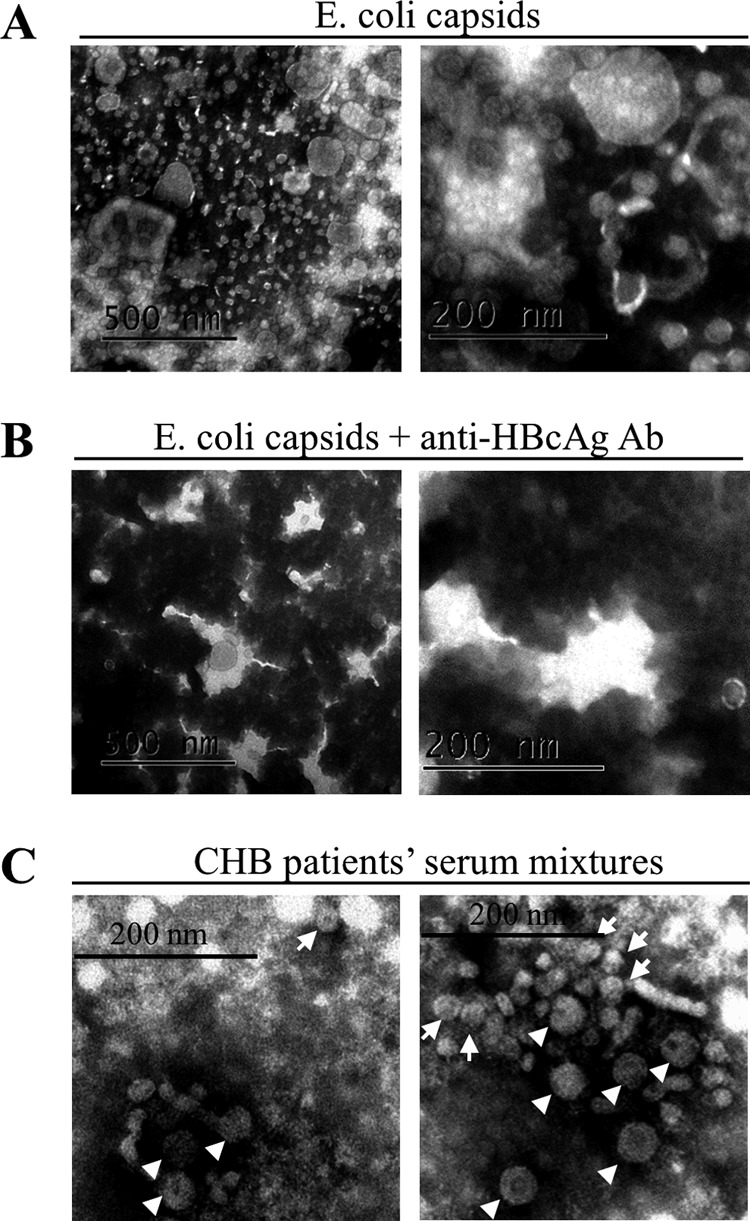
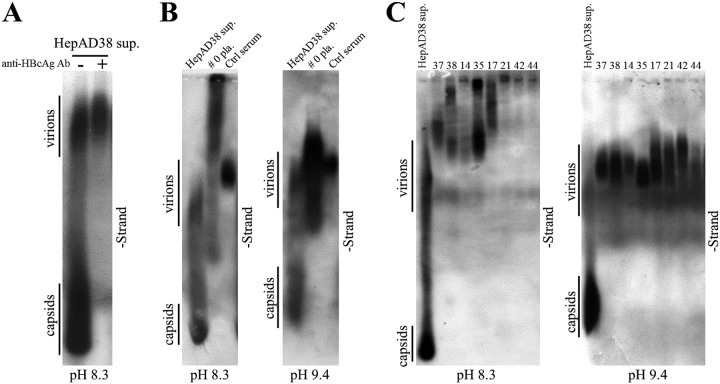
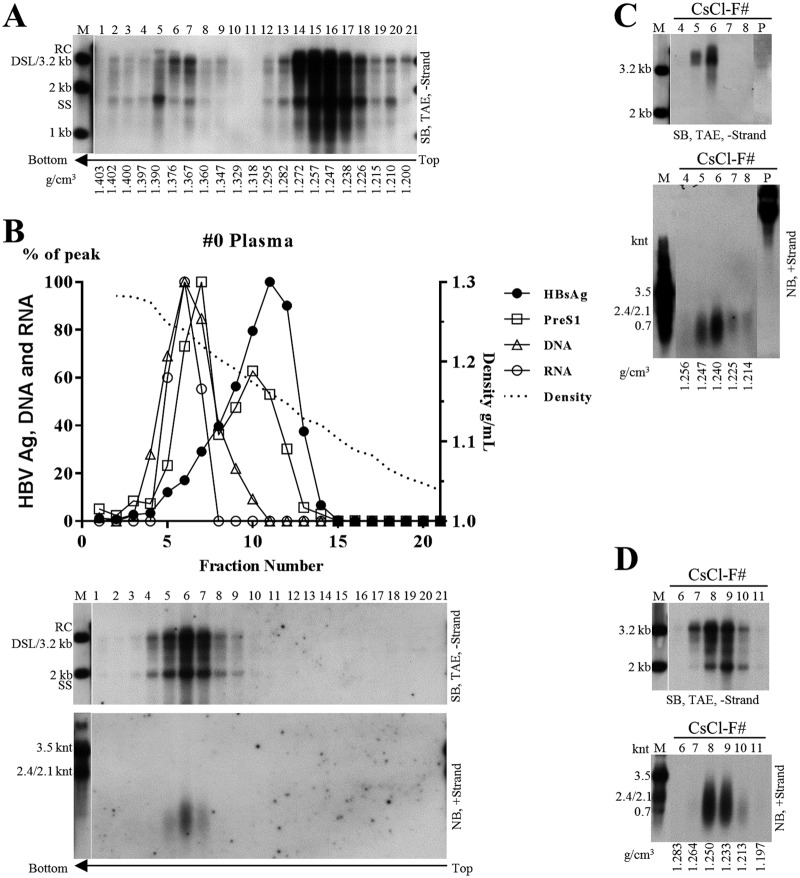
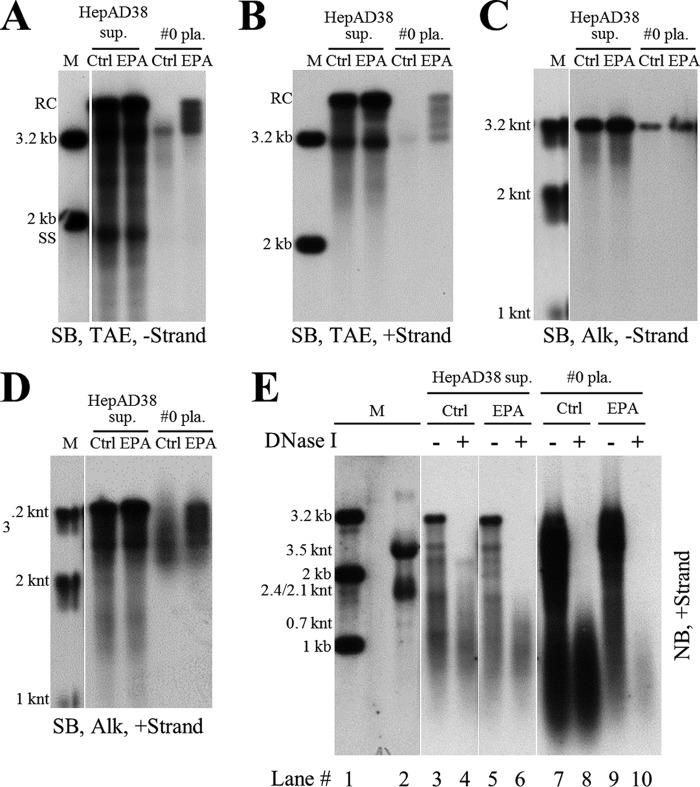
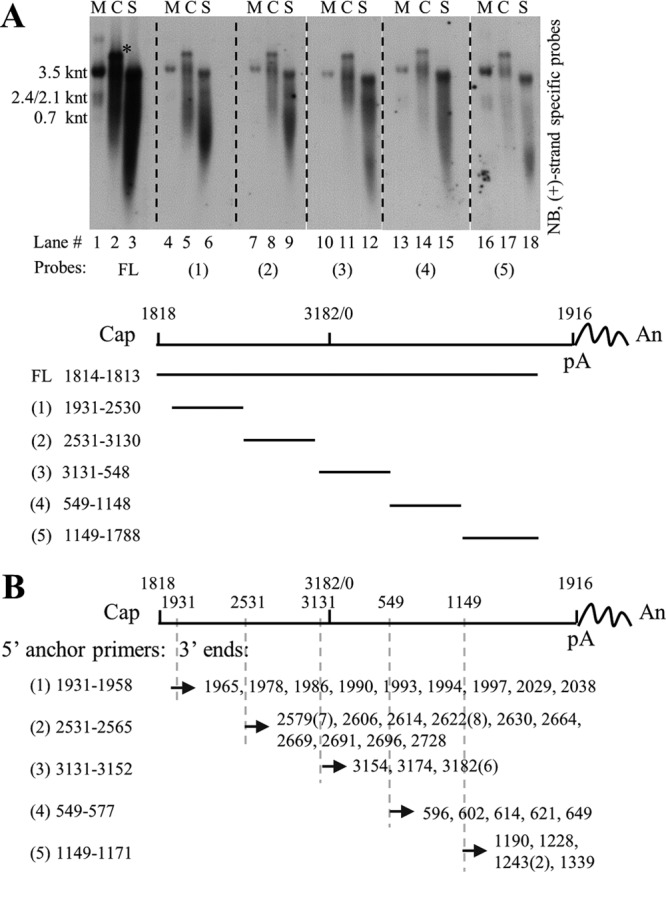
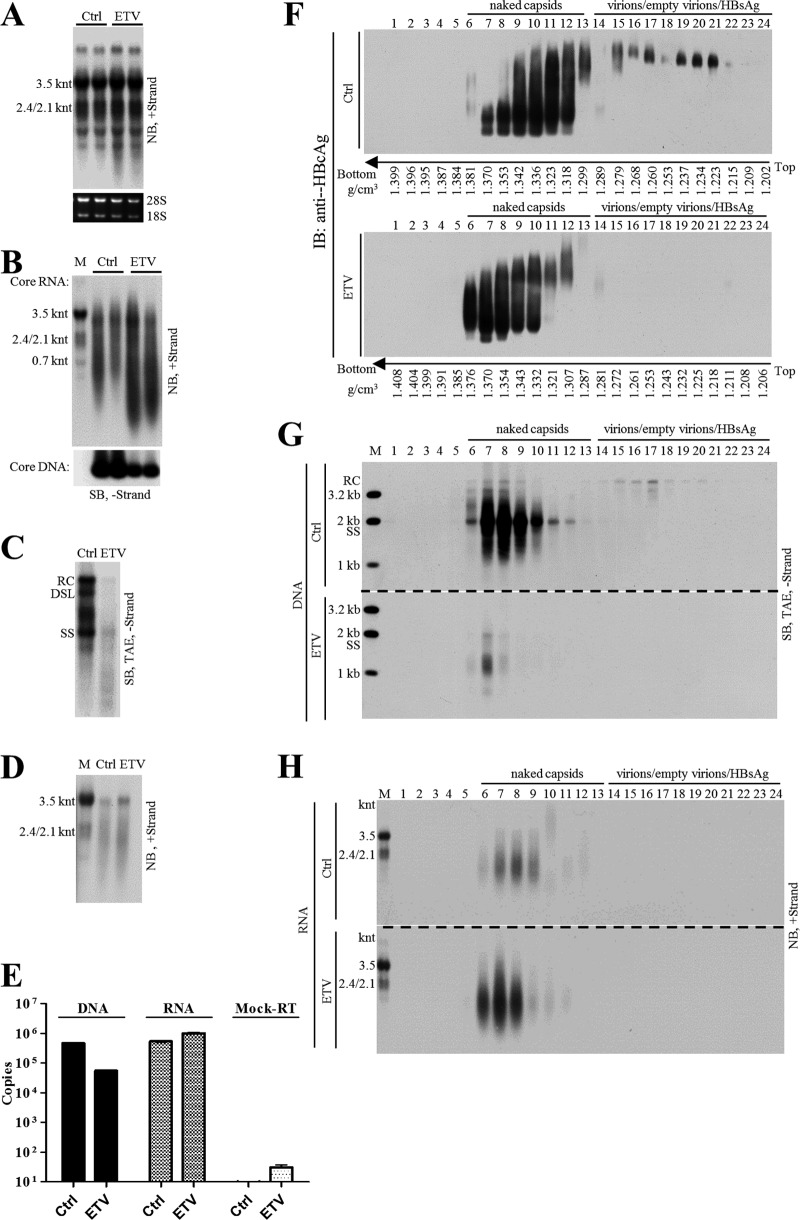
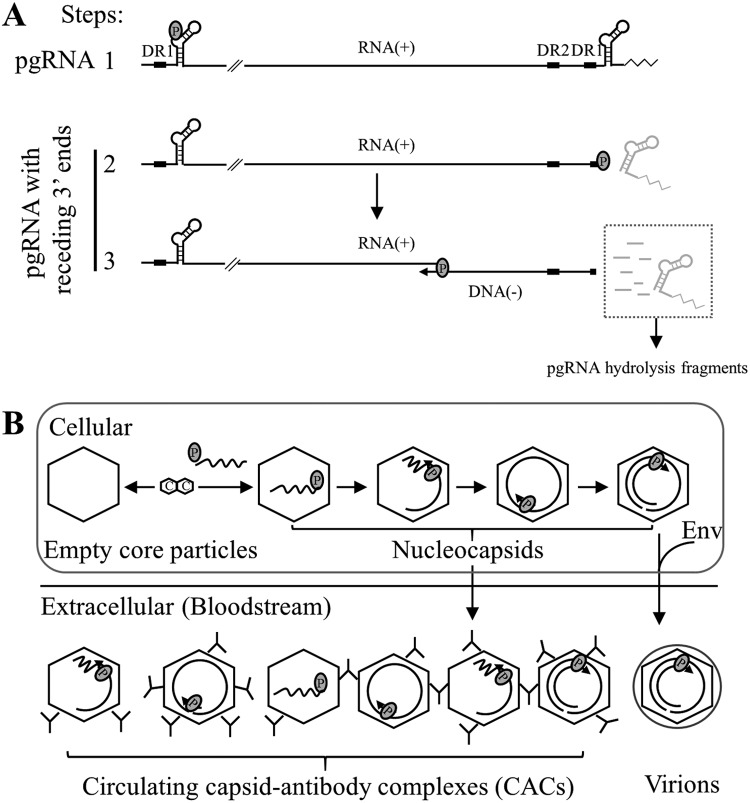
Extracellular HBV RNA has been detected in both HBV-replicating cell culture media and sera from chronic hepatitis B (CHB) patients, but its exact origin and composition remain controversial. Here, we demonstrated that extracellular HBV RNA species were of heterogeneous lengths, ranging from the length of pregenomic RNA to a few hundred nucleotides. In cell models, these RNAs were predominantly associated with naked capsids, although virions also harbored a minority of them. Moreover, HBV RNAs in hepatitis B patients' blood circulation were localized in unenveloped capsids in the form of capsid-antibody complexes (CACs) and in virions. Furthermore, we showed that extracellular HBV RNAs could serve as the template for viral DNA synthesis. In conclusion, extracellular HBV RNAs mainly consist of pgRNA or the pgRNA species degraded by the RNase H domain of the polymerase in the process of viral DNA synthesis and circulate as CACs and virions. Their presence in blood circulation of CHB patients may be exploited to develop novel biomarkers for HBV persistence.IMPORTANCE Although increasing evidence suggests the presence of extracellular HBV RNA species, their origin and molecular forms are still under debate. In addition to the infectious virions, HBV is known to secrete several species of incomplete viral particles, including hepatitis B surface antigen (HBsAg) particles, naked capsids, and empty virions, during its replication cycle. Here, we demonstrated that extracellular HBV RNAs were associated with naked capsids and virions in HepAD38 cells. Interestingly, we found that unenveloped capsids circulate in the blood of hepatitis B patients in the form of CACs and, together with virions, serve as vehicles carrying these RNA molecules. Moreover, extracellular HBV RNAs are heterogeneous in length and represent either pregenomic RNA (pgRNA) or products of incomplete reverse transcription during viral replication. These findings provide a conceptual basis for further application of extracellular RNA species as novel biomarkers for HBV persistence.
Keywords: capsid; capsid-antibody complex; extracellular HBV RNA; hepatitis B virus.
Copyright © 2018 Bai et al.
Figures
References
-
- Seeger C, Zoulim F, Mason WS. 2007. Hepadnaviruses, p 2977 In Knipe DM, Howley PM (ed), Fields virology, 5th ed, vol 2, Lippincott, Williams & Wilkins, Philadelphia.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
