

Color statistics of objects, and color tuning of object cortex in macaque monkey
- PMID: 30285103
- PMCID: PMC6168048
- DOI: 10.1167/18.11.1
Color statistics of objects, and color tuning of object cortex in macaque monkey
Abstract
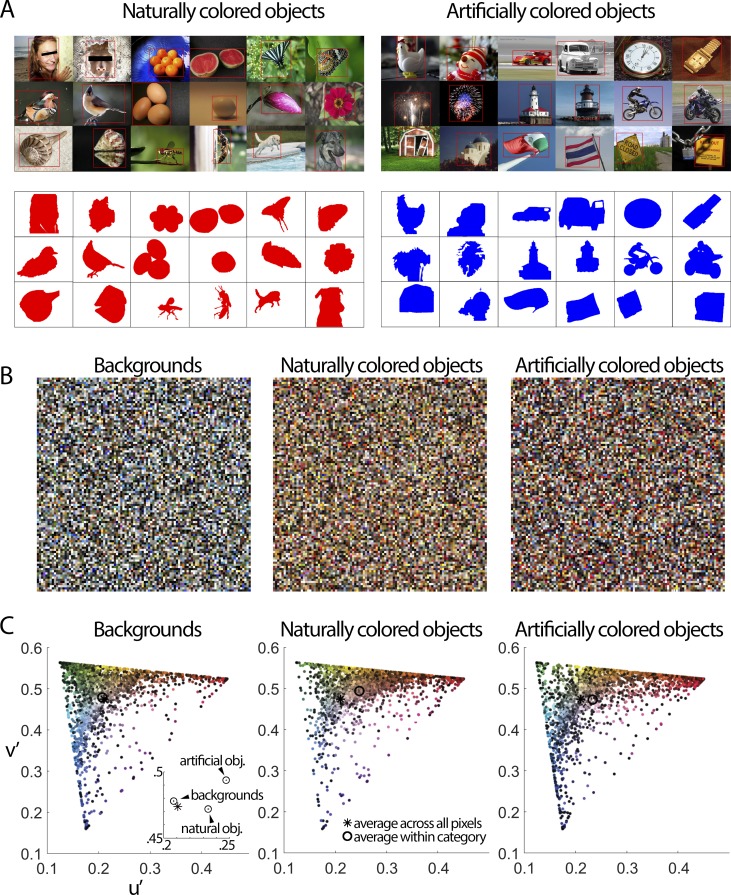
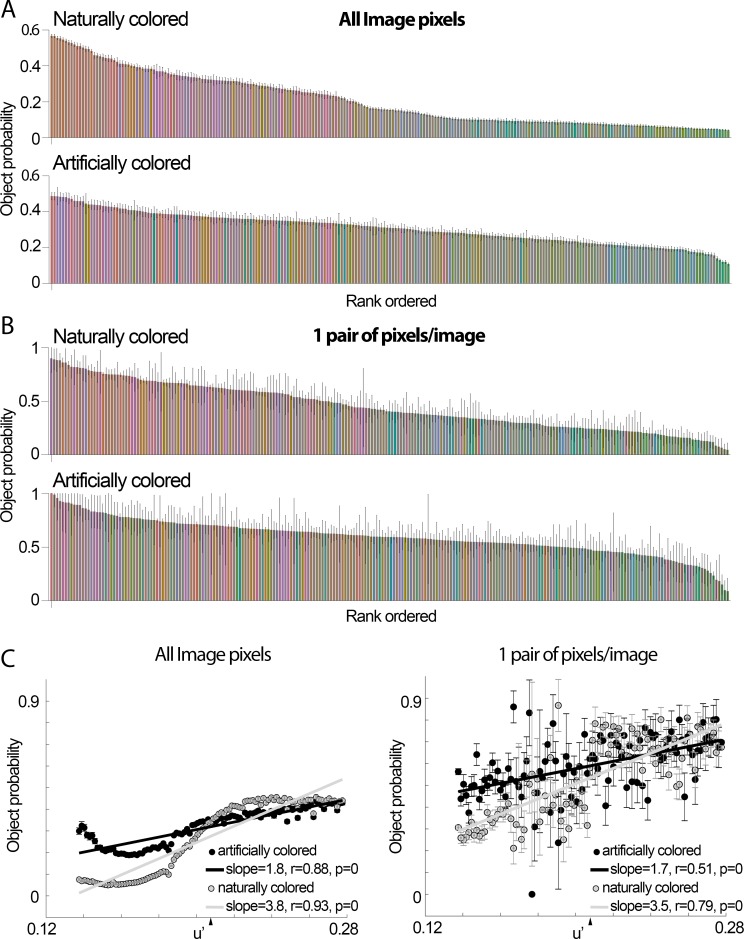
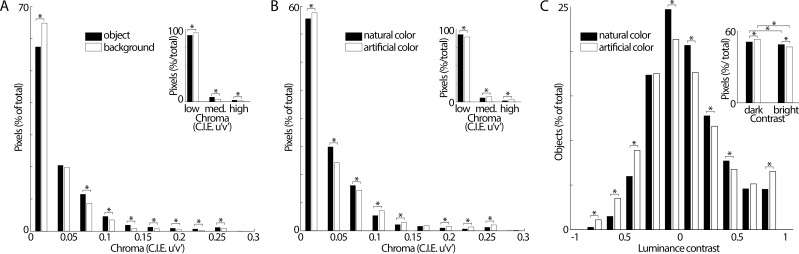
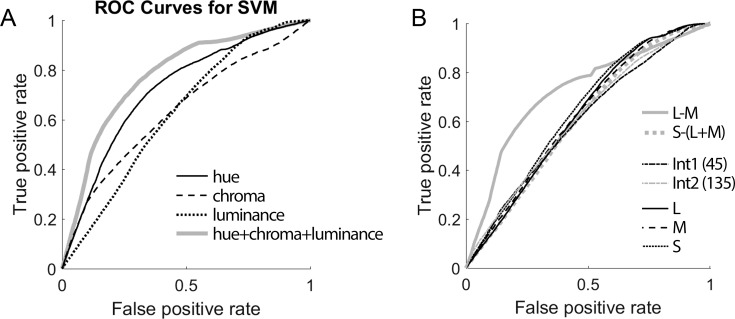
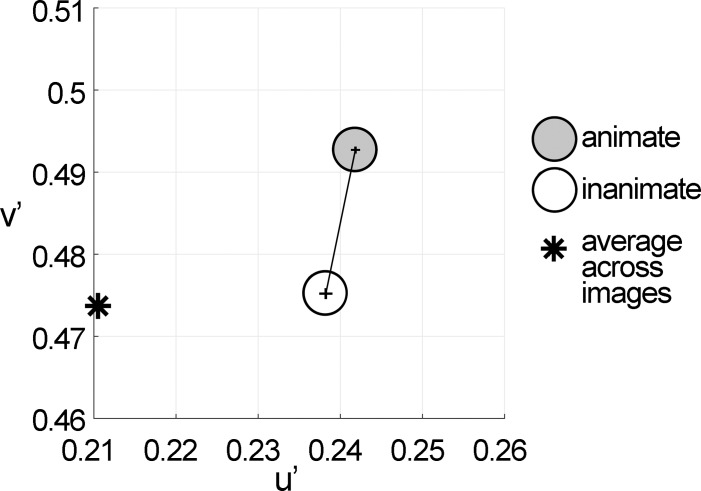
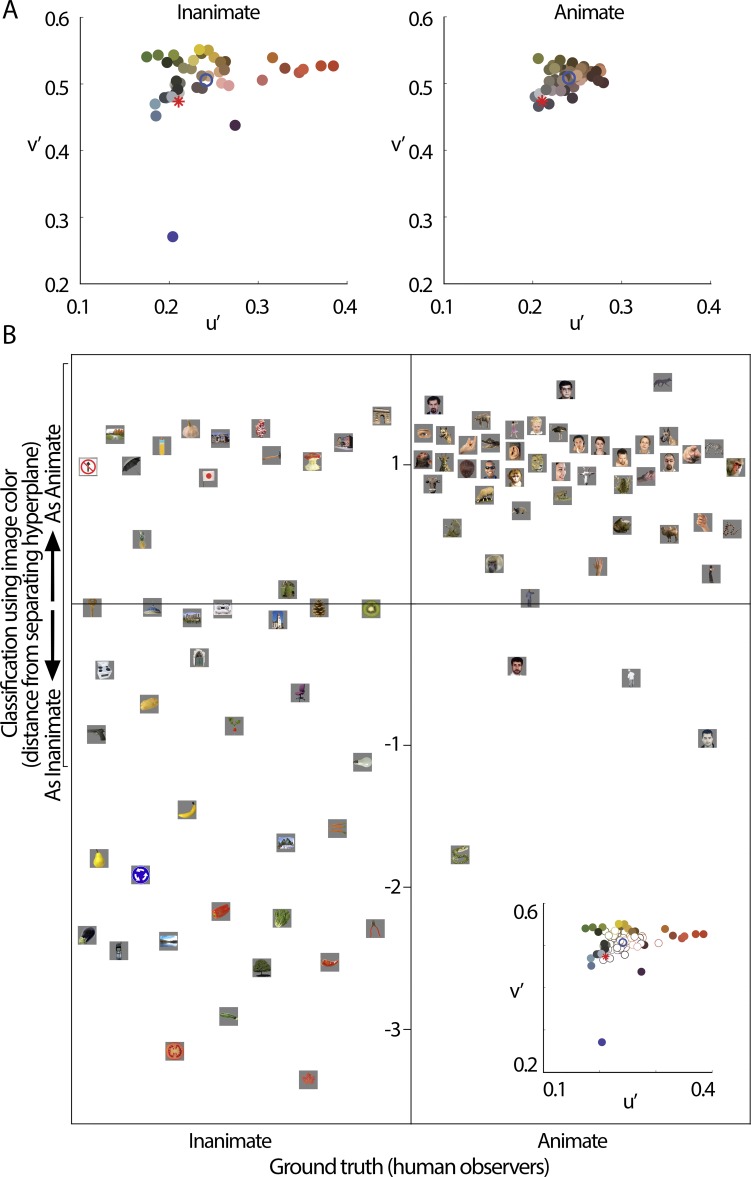
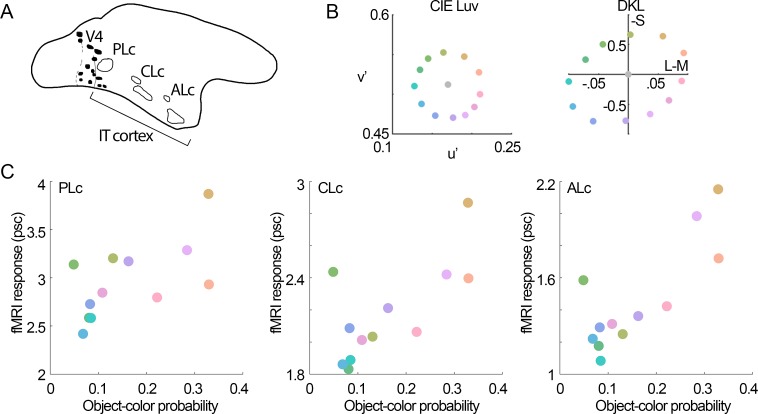
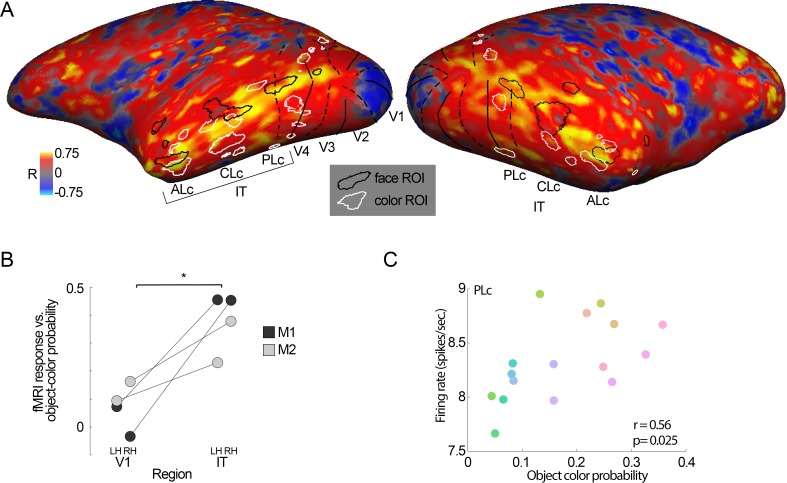
We hypothesized that the parts of scenes identified by human observers as "objects" show distinct color properties from backgrounds, and that the brain uses this information towards object recognition. To test this hypothesis, we examined the color statistics of naturally and artificially colored objects and backgrounds in a database of over 20,000 images annotated with object labels. Objects tended to be warmer colored (L-cone response > M-cone response) and more saturated compared to backgrounds. That the distinguishing chromatic property of objects was defined mostly by the L-M post-receptoral mechanism, rather than the S mechanism, is consistent with the idea that trichromatic color vision evolved in response to a selective pressure to identify objects. We also show that classifiers trained using only color information could distinguish animate versus inanimate objects, and at a performance level that was comparable to classification using shape features. Animate/inanimate is considered a fundamental superordinate category distinction, previously thought to be computed by the brain using only shape information. Our results show that color could contribute to animate/inanimate, and likely other, object-category assignments. Finally, color-tuning measured in two macaque monkeys with functional magnetic resonance imaging (fMRI), and confirmed by fMRI-guided microelectrode recording, supports the idea that responsiveness to color reflects the global functional organization of inferior temporal cortex, the brain region implicated in object vision. More strongly in IT than in V1, colors associated with objects elicited higher responses than colors less often associated with objects.
Figures
Similar articles
-
Disentangling Representations of Object Shape and Object Category in Human Visual Cortex: The Animate-Inanimate Distinction.J Cogn Neurosci. 2016 May;28(5):680-92. doi: 10.1162/jocn_a_00924. Epub 2016 Jan 14. J Cogn Neurosci. 2016. PMID: 26765944
-
Cortical representation of animate and inanimate objects in complex natural scenes.J Physiol Paris. 2012 Sep-Dec;106(5-6):239-49. doi: 10.1016/j.jphysparis.2012.02.001. Epub 2012 Mar 28. J Physiol Paris. 2012. PMID: 22472178 Free PMC article.
-
Stimulus representations in body-selective regions of the macaque cortex assessed with event-related fMRI.Neuroimage. 2012 Nov 1;63(2):723-41. doi: 10.1016/j.neuroimage.2012.07.013. Epub 2012 Jul 14. Neuroimage. 2012. PMID: 22796995
-
Color signals through dorsal and ventral visual pathways.Vis Neurosci. 2014 Mar;31(2):197-209. doi: 10.1017/S0952523813000382. Epub 2013 Oct 8. Vis Neurosci. 2014. PMID: 24103417 Free PMC article. Review.
-
Signals Related to Color in the Early Visual Cortex.Annu Rev Vis Sci. 2020 Sep 15;6:287-311. doi: 10.1146/annurev-vision-121219-081801. Annu Rev Vis Sci. 2020. PMID: 32936735 Review.
Cited by
-
Recent advances in understanding object recognition in the human brain: deep neural networks, temporal dynamics, and context.F1000Res. 2020 Jun 11;9:F1000 Faculty Rev-590. doi: 10.12688/f1000research.22296.1. eCollection 2020. F1000Res. 2020. PMID: 32566136 Free PMC article. Review.
-
Emergent color categorization in a neural network trained for object recognition.Elife. 2022 Dec 13;11:e76472. doi: 10.7554/eLife.76472. Elife. 2022. PMID: 36511778 Free PMC article.
-
Communication efficiency of color naming across languages provides a new framework for the evolution of color terms.Cognition. 2020 Feb;195:104086. doi: 10.1016/j.cognition.2019.104086. Epub 2019 Nov 12. Cognition. 2020. PMID: 31731116 Free PMC article.
-
Color appearance and the end of Hering's Opponent-Colors Theory.Trends Cogn Sci. 2023 Sep;27(9):791-804. doi: 10.1016/j.tics.2023.06.003. Epub 2023 Jul 1. Trends Cogn Sci. 2023. PMID: 37394292 Free PMC article. Review.
-
Color Tuning of Face-Selective Neurons in Macaque Inferior Temporal Cortex.eNeuro. 2021 Apr 12;8(2):ENEURO.0395-20.2020. doi: 10.1523/ENEURO.0395-20.2020. Print 2021 Mar-Apr. eNeuro. 2021. PMID: 33483324 Free PMC article.
References
-
- Bartels A, Zeki S. The architecture of the colour centre in the human visual brain: New results and a review. The European Journal of Neuroscience. (2000);12(1):172–193. - PubMed
-
- Beauchamp M. S, Haxby J. V, Jennings J. E, DeYoe E. A. An fMRI version of the Farnsworth-Munsell 100-Hue test reveals multiple color-selective areas in human ventral occipitotemporal cortex. Cerebral Cortex. (1999);9(3):257–263. - PubMed
-
- Caramazza A, Shelton J. R. Domain-specific knowledge systems in the brain: The animate-inanimate distinction. Journal of Cognitive Neuroscience. (1998);10(1):1–34. - PubMed
-
- Chang C.-C, Lin C.-J. LIBSVM : A library for support vector machines. ACM Transactions on Intelligent Systems and Technology. (2011);2(27):1–27. http://www.csie.ntu.edu.tw/∼cjlin/libsvm.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
