

Molecular characterization of three novel perforins in common carp (Cyprinus carpio L.) and their expression patterns during larvae ontogeny and in response to immune challenges
- PMID: 30285759
- PMCID: PMC6169072
- DOI: 10.1186/s12917-018-1613-y
Molecular characterization of three novel perforins in common carp (Cyprinus carpio L.) and their expression patterns during larvae ontogeny and in response to immune challenges
Abstract
Background: In the host immune system, perforin is a cytotoxic effector molecule that eliminate virus-infected and malignant cells. Moreover, some recent studies also imply the involvement of perforin in antibacterial immunity. Common carp (Cyprinus carpio L.), one of the most economically important fish species in China, has a high susceptibility to viruses and bacteria. Thus far, in common carp, no data are available regarding the identification and immunologic function of the perforin.
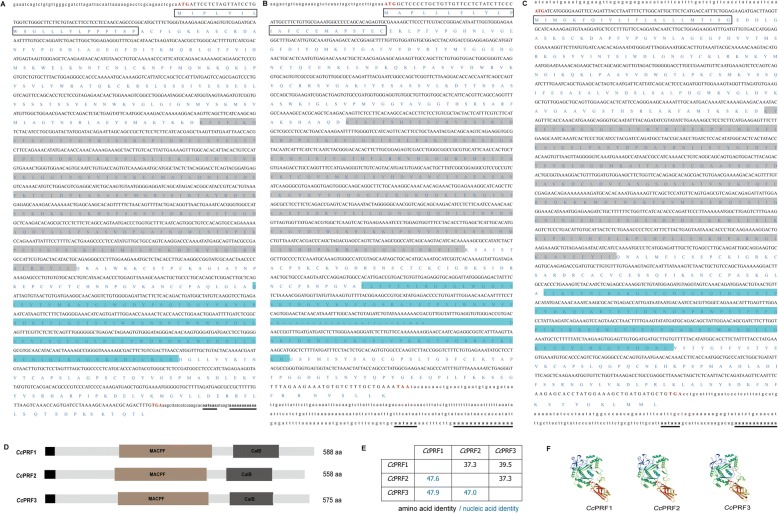
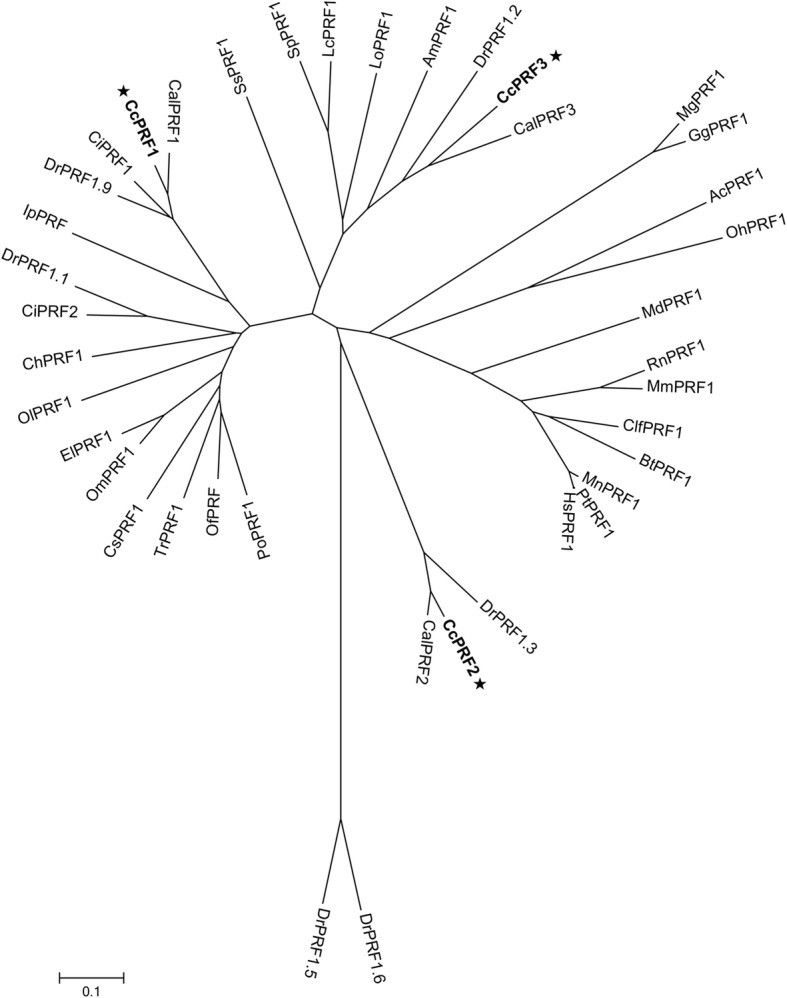
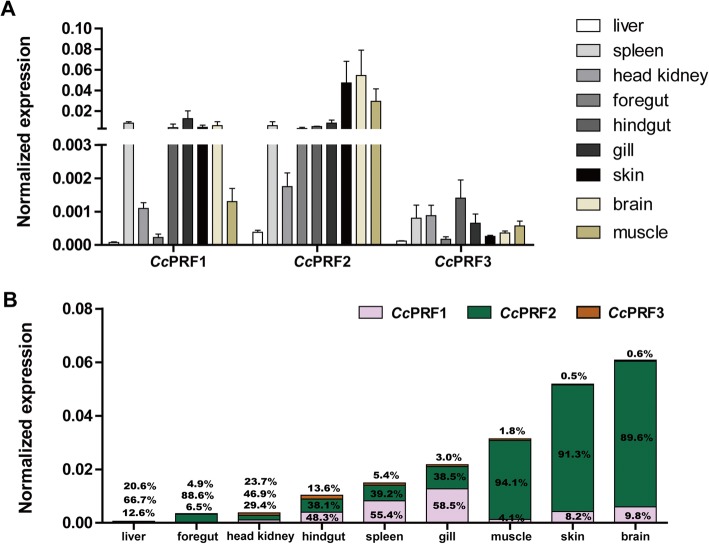
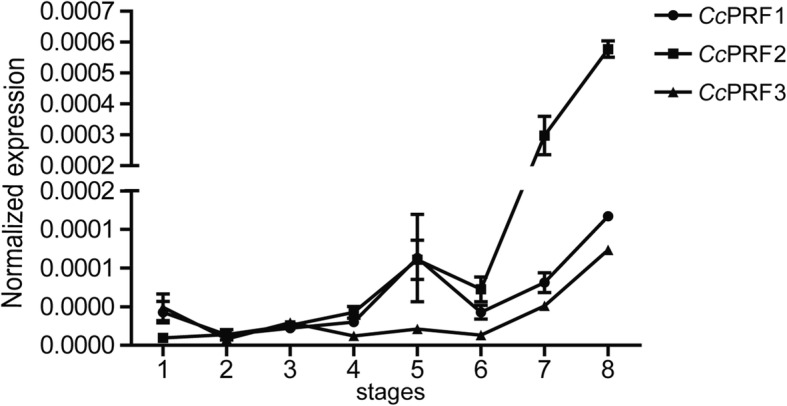
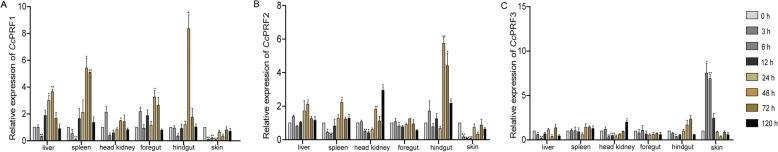
Results: In the present study, the cDNA and genomic DNA sequences of three perforin isoform genes were cloned and characterized in common carp, named CcPRF1, CcPRF2 and CcPRF3. Amino acid sequences of the three CcPRFs were quite different, with identities ranged from 37.3 to 39.5%. Phylogenetic analysis showed that three CcPRFs, each in a separate sub-branch, possessed closer evolutionary relationship with other teleost perforins, especially with cyprinid fishes, than higher vertebrates. Expression analysis revealed that each CcPRF gene was differentially expressed in all of the nine tested tissues. During larvae ontogeny, each CcPRF displayed a distinct expression pattern, while with a common expression peak at 22 days post hatching (dph). Moreover, in vivo or in vitro, after stimulation with polyI:C, LPS and Aeromonas hydrophila, each CcPRF was induced significantly, with differential expression dynamics.
Conclusions: Our findings suggest that perforin might play significant roles in larval immune system and in the immune defense of common carp against viral and bacterial pathogens. Meantime, the differential expression dynamics seem to imply possible different cellular locations or functional differences across various CcPRF isoforms.
Keywords: Common carp (Cyprinus carpio L.); Evolutionary relationship; Expression pattern; Perforin.
Conflict of interest statement
Ethics approval and consent to participate
The protocol was approved by the Animal Experimental Ethics Committee of Shandong Normal University (Permit Number: AEECSDNU2017005). Additionally, the ARRIVE (Animal Research: Reporting of in Vivo Experiments) checklist has been displayed in Additional file 10.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures



References
-
- Tokmadzic VS, Tomas MI, Sotosek S, Laskarin G, Dominovic M, Tulic V, Dordevic G, Sustic A, Mrakovcic-Sutic I. Different perforin expression in peripheral blood and prostate tissue in patients with benign prostatic hyperplasia and prostate cancer. Scand J Immunol. 2011;74(4):368–376. doi: 10.1111/j.1365-3083.2011.02569.x. - DOI - PubMed
-
- Alexander AA, Maniar A, Cummings JS, Hebbeler AM, Schulze DH, Gastman BR, Pauza CD, Strome SE, Chapoval AI. Isopentenyl pyrophosphate-activated CD56+ {gamma}{delta} T lymphocytes display potent antitumor activity toward human squamous cell carcinoma. Clin Cancer Res. 2008;14(13):4232–4240. doi: 10.1158/1078-0432.CCR-07-4912. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
