

Reduced menin expression impairs rapamycin effects as evidenced by an increase in mTORC2 signaling and cell migration
- PMID: 30285764
- PMCID: PMC6167842
- DOI: 10.1186/s12964-018-0278-2
Reduced menin expression impairs rapamycin effects as evidenced by an increase in mTORC2 signaling and cell migration
Abstract
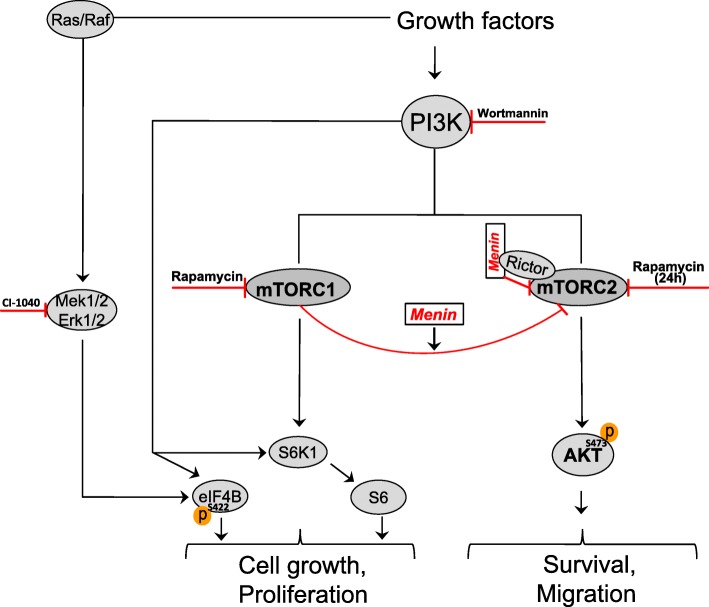
Background: Mammalian target of rapamycin (mTOR) is a master regulator of various cellular responses by forming two functional complexes, mTORC1 and mTORC2. mTOR signaling is frequently dysregulated in pancreatic neuroendocrine tumors (PNETs). mTOR inhibitors have been used in attempts to treat these lesions, and prolonged progression free survival has been recorded. If this holds true also for the multiple endocrine neoplasia type 1 (MEN1) associated PNETs is yet unclear. We investigated the relationship between expression of the MEN1 protein menin and mTOR signaling in the presence or absence of the mTOR inhibitor rapamycin.
Methods: In addition to use of menin wild type and menin-null mouse embryonic fibroblasts (MEFs), menin was silenced by siRNA in pancreatic neuroendocrine tumor cell line BON-1. Panels of protein phosphorylation, as activation markers downstream of PI3k-mTOR-Akt pathways, as well as menin expression were evaluated by immunoblotting. The impact of menin expression in the presence and absence of rapamycin was determinate upon Wound healing, migration and proliferation in MEFs and BON1 cells.
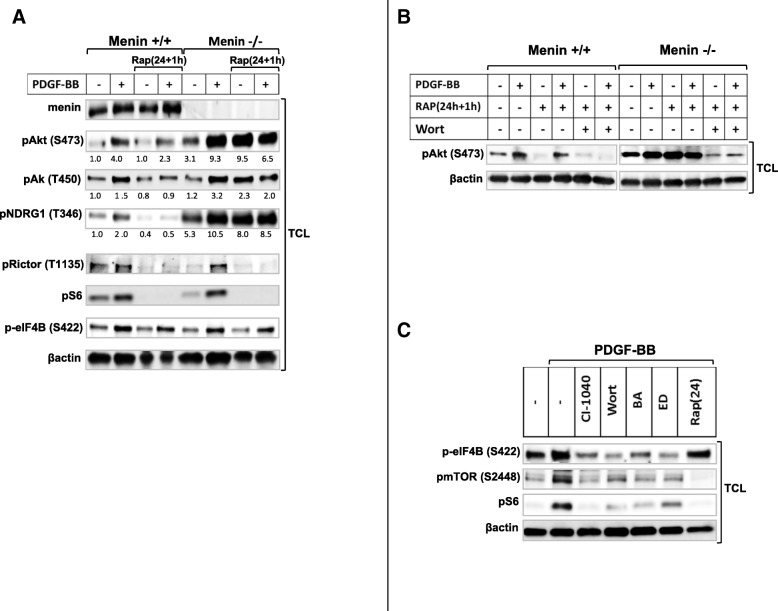
Results: PDGF-BB markedly increased phosphorylation of mTORC2 substrate Akt, at serine 473 (S473) and threonine 450 (T450) in menin-/- MEFs but did not alter phosphorylation of mTORC1 substrates ribosomal protein S6 or eIF4B. Acute rapamycin treatment by mTORC1-S6 inhibition caused a greater enhancement of Akt phosphorylation on S473 in menin-/- cells as compared to menin+/+ MEFs (116% vs 38%). Chronic rapamycin treatment, which inhibits both mTORC1and 2, reduced Akt phosphorylation of S473 to a lesser extent in menin-/- MEFs than menin+/+ MEFs (25% vs 75%). Silencing of menin expression in human PNET cell line (BON1) also enhanced Akt phosphorylation at S473, but not activation of mTORC1. Interestingly, silencing menin in BON1 cells elevated S473 phosphorylation of Akt in both acute and chronic treatments with rapamycin. Finally, we show that the inhibitory effect of rapamycin on serum mediated wound healing and cell migration is impaired in menin-/- MEFs, as well as in menin-silenced BON1 cells.
Conclusions: Menin is involved in regulatory mechanism between the two mTOR complexes, and its reduced expression is accompanied with increased mTORC2-Akt signaling, which consequently impairs anti-migratory effect of rapamycin.
Keywords: Akt; MEN1; PI3K; PNET; Rapamycin; mTOR.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures



Similar articles
-
Involvement of mTORC1 and mTORC2 in regulation of glioblastoma multiforme growth and motility.Int J Oncol. 2009 Oct;35(4):731-40. doi: 10.3892/ijo_00000386. Int J Oncol. 2009. PMID: 19724909
-
Increased AKT S473 phosphorylation after mTORC1 inhibition is rictor dependent and does not predict tumor cell response to PI3K/mTOR inhibition.Mol Cancer Ther. 2009 Apr;8(4):742-53. doi: 10.1158/1535-7163.MCT-08-0668. Mol Cancer Ther. 2009. PMID: 19372546 Free PMC article.
-
Diverse signaling mechanisms of mTOR complexes: mTORC1 and mTORC2 in forming a formidable relationship.Adv Biol Regul. 2019 May;72:51-62. doi: 10.1016/j.jbior.2019.03.003. Epub 2019 Apr 11. Adv Biol Regul. 2019. PMID: 31010692 Review.
-
Discrete signaling mechanisms of mTORC1 and mTORC2: Connected yet apart in cellular and molecular aspects.Adv Biol Regul. 2017 May;64:39-48. doi: 10.1016/j.jbior.2016.12.001. Epub 2017 Jan 4. Adv Biol Regul. 2017. PMID: 28189457 Review.
-
Active-site inhibitors of mTOR target rapamycin-resistant outputs of mTORC1 and mTORC2.PLoS Biol. 2009 Feb 10;7(2):e38. doi: 10.1371/journal.pbio.1000038. PLoS Biol. 2009. PMID: 19209957 Free PMC article.
Cited by
-
CDK7 blockade suppresses super-enhancer-associated oncogenes in bladder cancer.Cell Oncol (Dordr). 2021 Aug;44(4):871-887. doi: 10.1007/s13402-021-00608-x. Epub 2021 Apr 27. Cell Oncol (Dordr). 2021. PMID: 33905040
-
Multiple endocrine neoplasia type 1 combined with thyroid neoplasm: A case report and review of literatures.World J Clin Cases. 2022 Jan 21;10(3):1032-1040. doi: 10.12998/wjcc.v10.i3.1032. World J Clin Cases. 2022. PMID: 35127917 Free PMC article.
-
Menin in Cancer.Genes (Basel). 2024 Sep 21;15(9):1231. doi: 10.3390/genes15091231. Genes (Basel). 2024. PMID: 39336822 Free PMC article. Review.
-
Generation and characterization of CRISPR/Cas9-mediated MEN1 knockout BON1 cells: a human pancreatic neuroendocrine cell line.Sci Rep. 2020 Sep 3;10(1):14572. doi: 10.1038/s41598-020-71516-7. Sci Rep. 2020. PMID: 32884006 Free PMC article.
-
The link between menin and pleiotrophin in the tumor biology of pancreatic neuroendocrine neoplasms.Cancer Sci. 2022 May;113(5):1575-1586. doi: 10.1111/cas.15301. Epub 2022 Mar 8. Cancer Sci. 2022. PMID: 35179814 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
