

Perfidious synaptic transmission in the guinea-pig auditory brainstem
- PMID: 30286113
- PMCID: PMC6172016
- DOI: 10.1371/journal.pone.0203712
Perfidious synaptic transmission in the guinea-pig auditory brainstem
Abstract
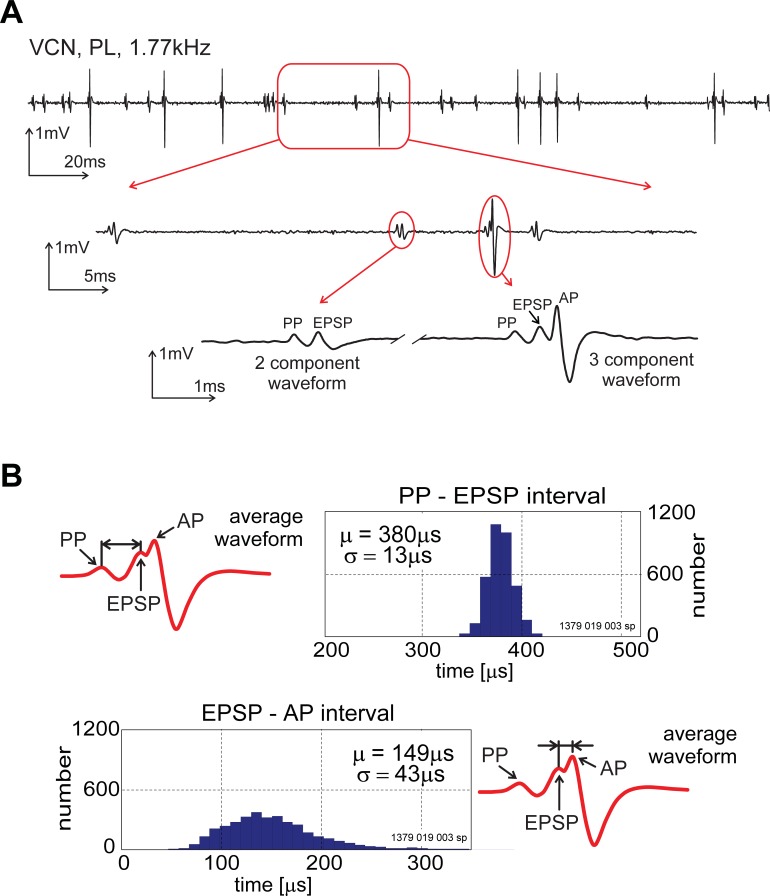
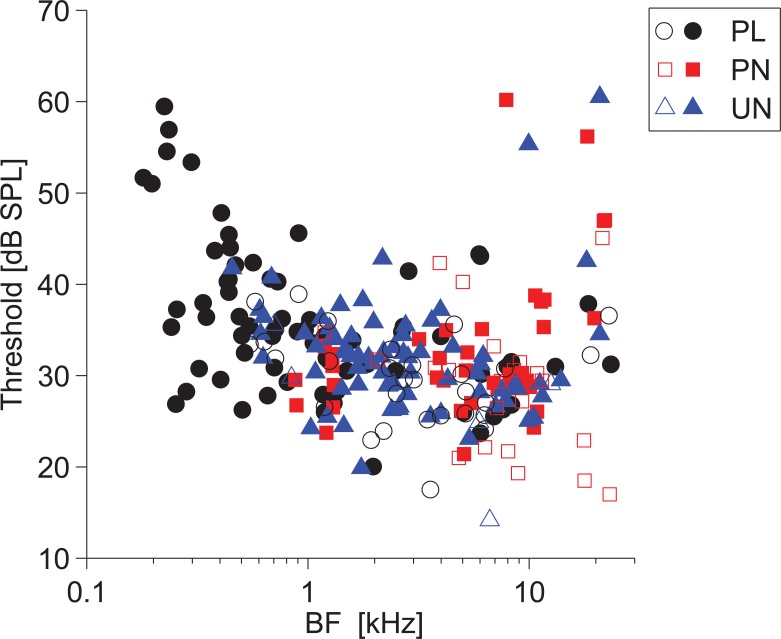
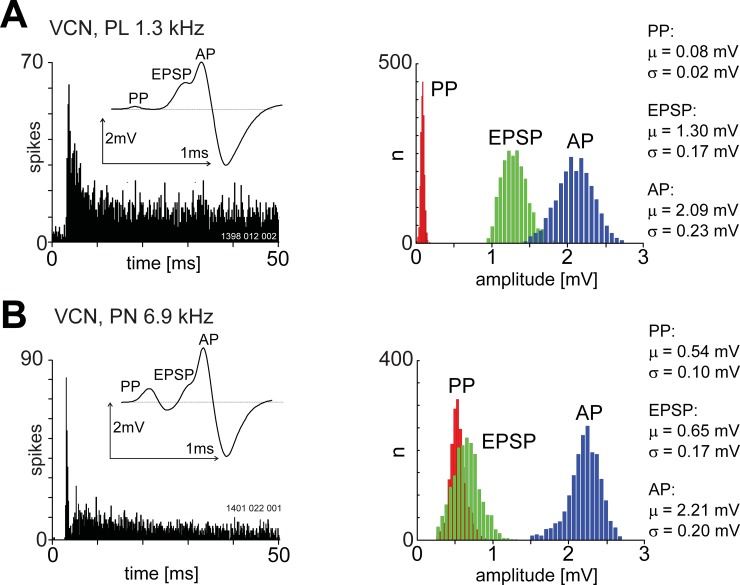
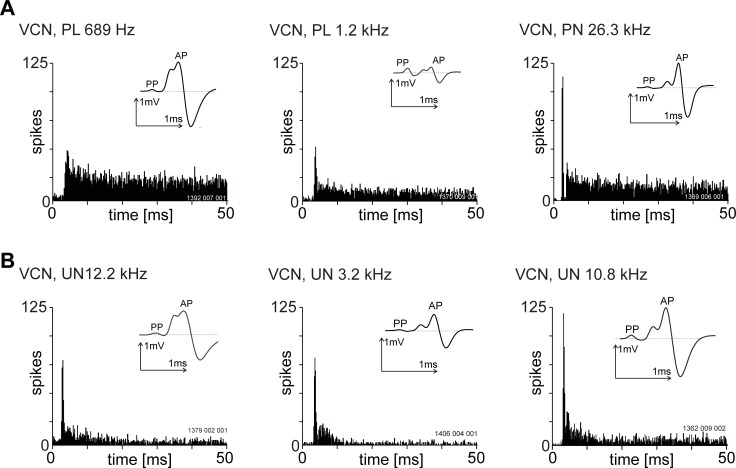
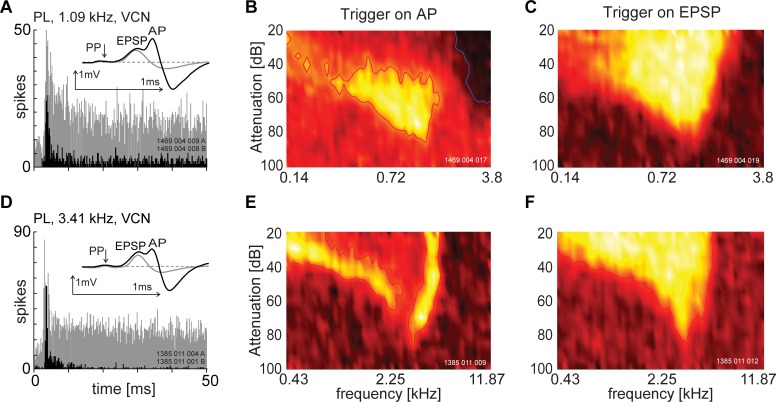
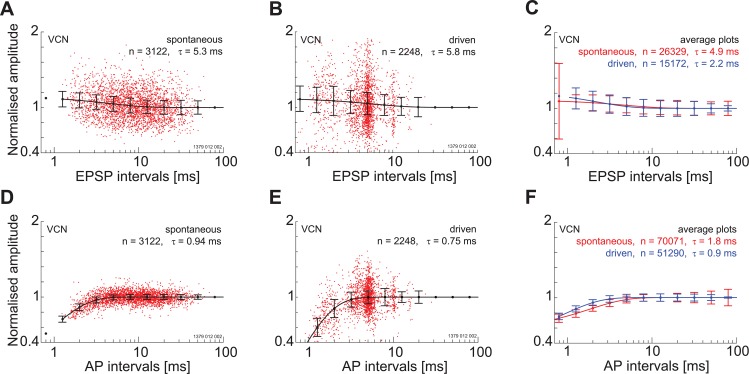
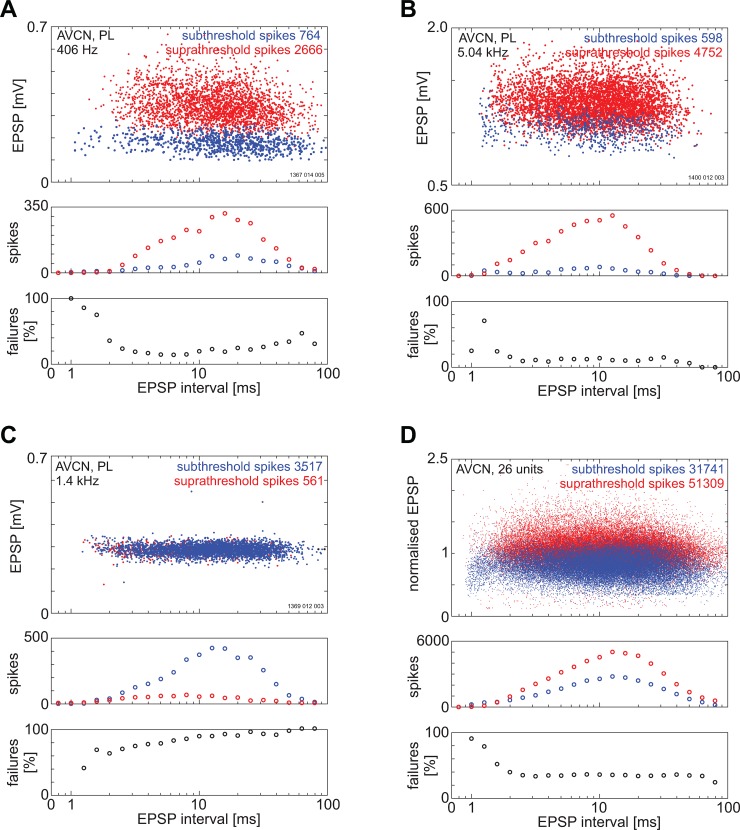
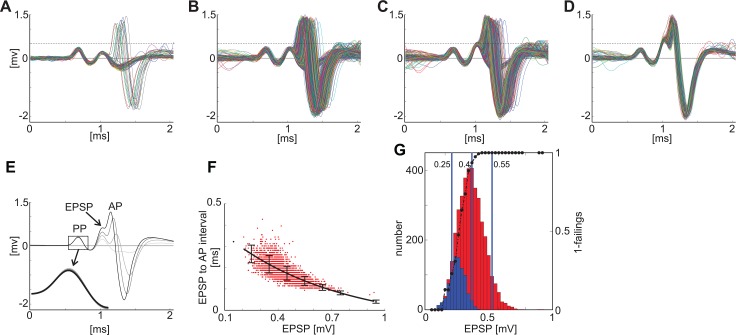
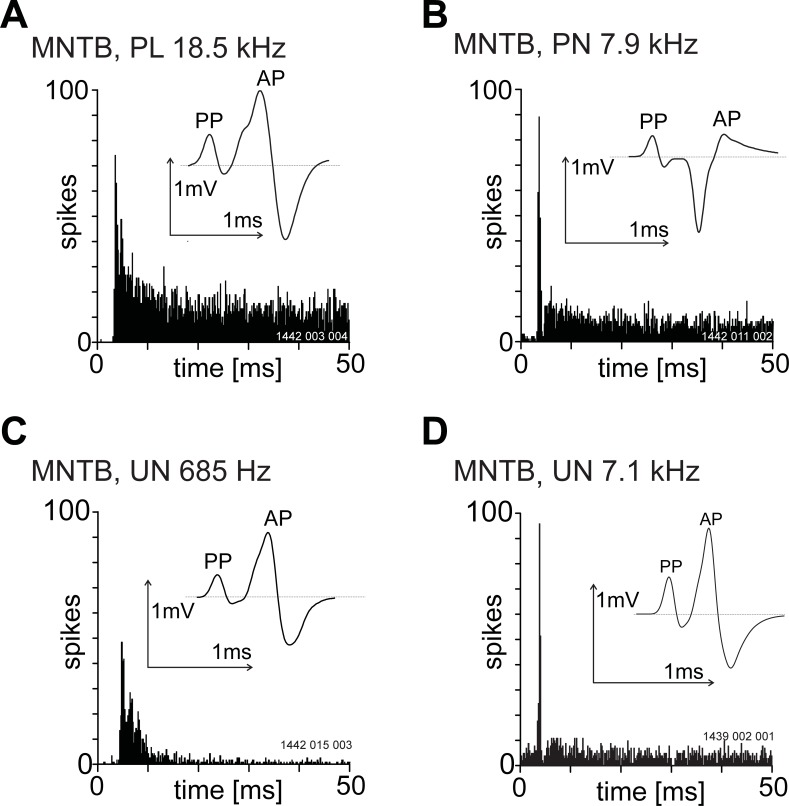
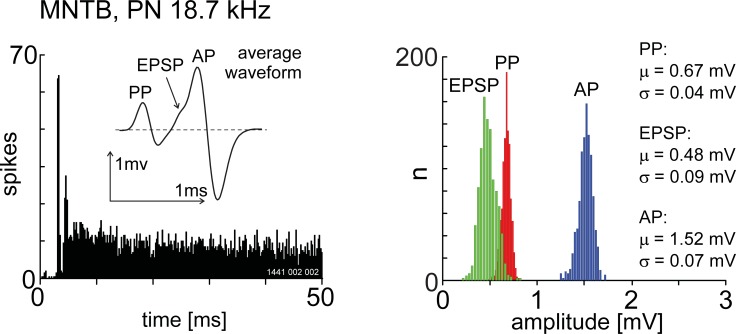
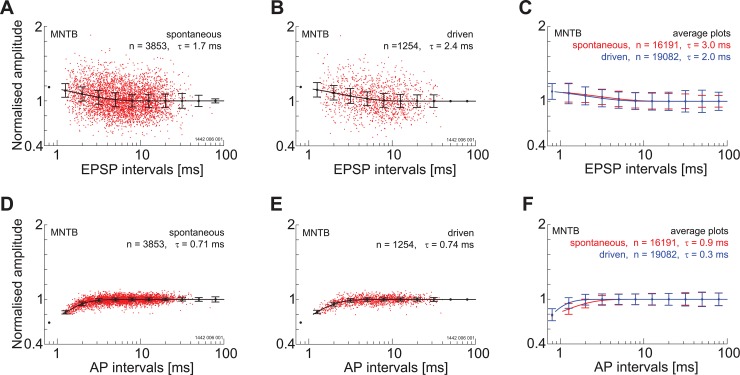
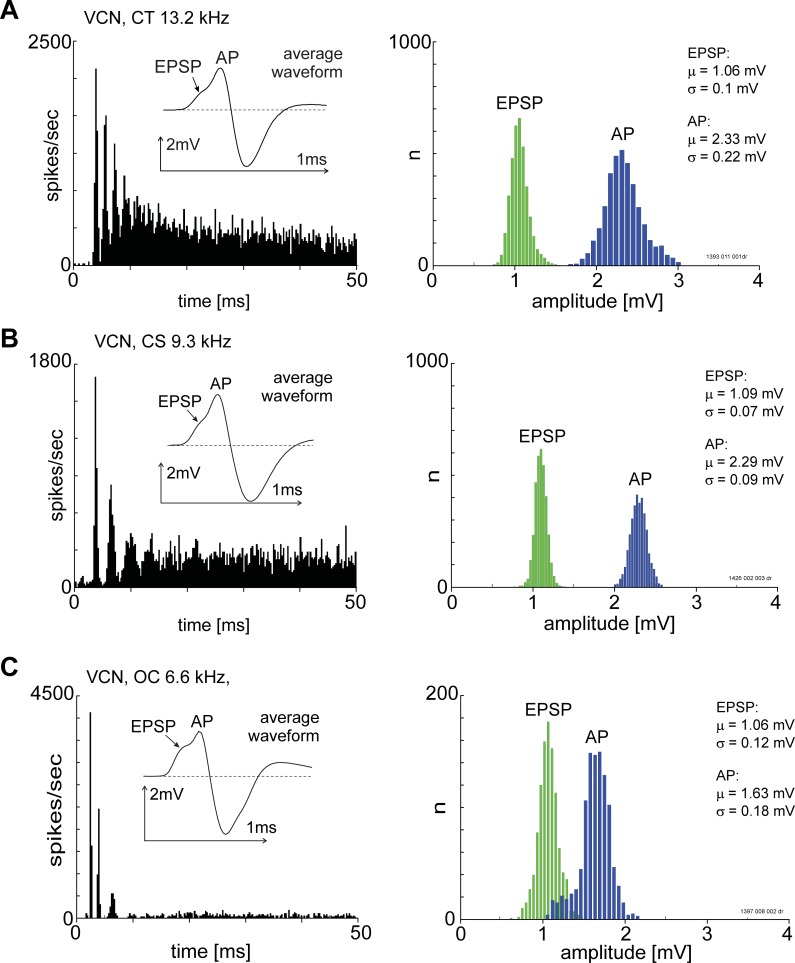
The presence of 'giant' synapses in the auditory brainstem is thought to be a specialization designed to encode temporal information to support perception of pitch, frequency, and sound-source localisation. These 'giant' synapses have been found in the ventral cochlear nucleus, the medial nucleus of the trapezoid body and the ventral nucleus of the lateral lemniscus. An interpretation of these synapses as simple relays has, however, been challenged by the observation in the gerbil that the action potential frequently fails in the ventral cochlear nucleus. Given the prominence of these synapses it is important to establish whether this phenomenon is unique to the gerbil or can be observed in other species. Here we examine the responses of units, thought to be the output of neurons in receipt of 'giant' synaptic endings, in the ventral cochlear nucleus and the medial nucleus of the trapezoid body in the guinea pig. We found that failure of the action-potential component, recorded from cells in the ventral cochlear nucleus, occurred in ~60% of spike waveforms when recording spontaneous activity. In the medial nucleus of the trapezoid body, we did not find evidence for action-potential failure. In the ventral cochlear nucleus action-potential failures transform the receptive field between input and output of bushy cells. Additionally, the action-potential failures result in "non-primary-like" temporal-adaptation patterns. This is important for computational models of the auditory system, which commonly assume the responses of ventral cochlear nucleus bushy cells are very similar to their "primary like" auditory-nerve-fibre inputs.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Synaptic Inhibition of Medial Olivocochlear Efferent Neurons by Neurons of the Medial Nucleus of the Trapezoid Body.J Neurosci. 2020 Jan 15;40(3):509-525. doi: 10.1523/JNEUROSCI.1288-19.2019. Epub 2019 Nov 12. J Neurosci. 2020. PMID: 31719165 Free PMC article.
-
Effect of altered neuronal activity on cell size in the medial nucleus of the trapezoid body and ventral cochlear nucleus of the gerbil.J Comp Neurol. 1994 Oct 1;348(1):111-20. doi: 10.1002/cne.903480106. J Comp Neurol. 1994. PMID: 7814680
-
Linear coding of complex sound spectra by discharge rate in neurons of the medial nucleus of the trapezoid body (MNTB) and its inputs.Front Neural Circuits. 2014 Dec 16;8:144. doi: 10.3389/fncir.2014.00144. eCollection 2014. Front Neural Circuits. 2014. PMID: 25565971 Free PMC article.
-
The multiple functions of T stellate/multipolar/chopper cells in the ventral cochlear nucleus.Hear Res. 2011 Jun;276(1-2):61-9. doi: 10.1016/j.heares.2010.10.018. Epub 2010 Nov 4. Hear Res. 2011. PMID: 21056098 Free PMC article. Review.
-
The ion channels and synapses responsible for the physiological diversity of mammalian lower brainstem auditory neurons.Hear Res. 2019 May;376:33-46. doi: 10.1016/j.heares.2018.12.011. Epub 2018 Dec 26. Hear Res. 2019. PMID: 30606624 Review.
Cited by
-
I' Wave Auditory Brainstem Response as a Possible Indicator of Noise-Induced Cochlear Synaptopathy.Iran J Child Neurol. 2025 Jun 25;19(3):77-82. doi: 10.22037/ijcn.v19i3.47308. eCollection 2025 Summer. Iran J Child Neurol. 2025. PMID: 40787274
-
Presynaptic Mitochondria Volume and Abundance Increase during Development of a High-Fidelity Synapse.J Neurosci. 2019 Oct 9;39(41):7994-8012. doi: 10.1523/JNEUROSCI.0363-19.2019. Epub 2019 Aug 27. J Neurosci. 2019. PMID: 31455662 Free PMC article.
-
Cellular and synaptic specializations for sub-millisecond precision in the mammalian auditory brainstem.Front Cell Neurosci. 2025 May 19;19:1568506. doi: 10.3389/fncel.2025.1568506. eCollection 2025. Front Cell Neurosci. 2025. PMID: 40458470 Free PMC article. Review.
-
Harmonic Cancellation-A Fundamental of Auditory Scene Analysis.Trends Hear. 2021 Jan-Dec;25:23312165211041422. doi: 10.1177/23312165211041422. Trends Hear. 2021. PMID: 34698574 Free PMC article.
References
-
- Borst JGG, Rusu SI. The Calyx of Held Synapse In: Trussell LO, Popper A, Fay RR, editors. Springer Handbook of Auditory Research: Synaptic Mechanisms in the Auditory System. New York: Springer; 2012. p. 95–134.
-
- Manis PB, Xie R, Wang Y, Marrs GS, Spirou GA. The Endbulbs of Held In: Trussell LO, Popper A, Fay RR, editors. Springer Handbook of Auditory Research: Synaptic Mechanisms in the Auditory System. New York: Springer; 2012. p. 61–94.
-
- Sayles M, Heinz MG. Afferent Coding and Efferent Control in the Normal and Impaired Cochlea In: Manley GA, Gummer AW, Popper AN, Fay RR, editors. Springer Handbook of Auditory Research: Understanding the Cochlea. 62 New York: Springer; 2017. p. 215–52.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
