

SOX4 controls invariant NKT cell differentiation by tuning TCR signaling
- PMID: 30287480
- PMCID: PMC6219734
- DOI: 10.1084/jem.20172021
SOX4 controls invariant NKT cell differentiation by tuning TCR signaling
Abstract
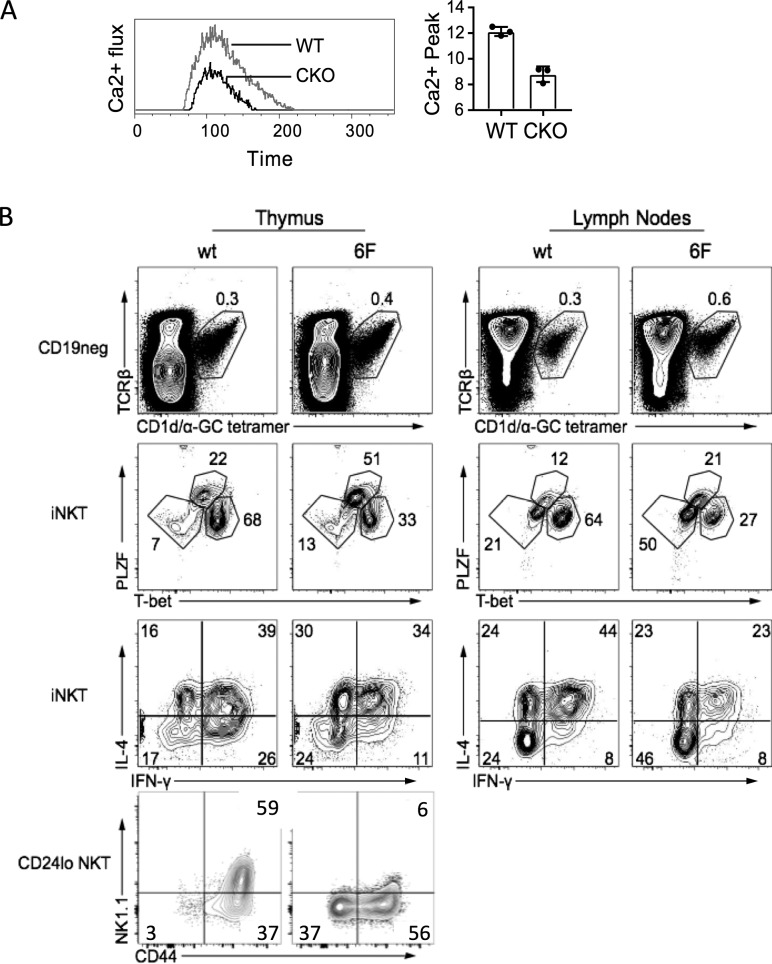
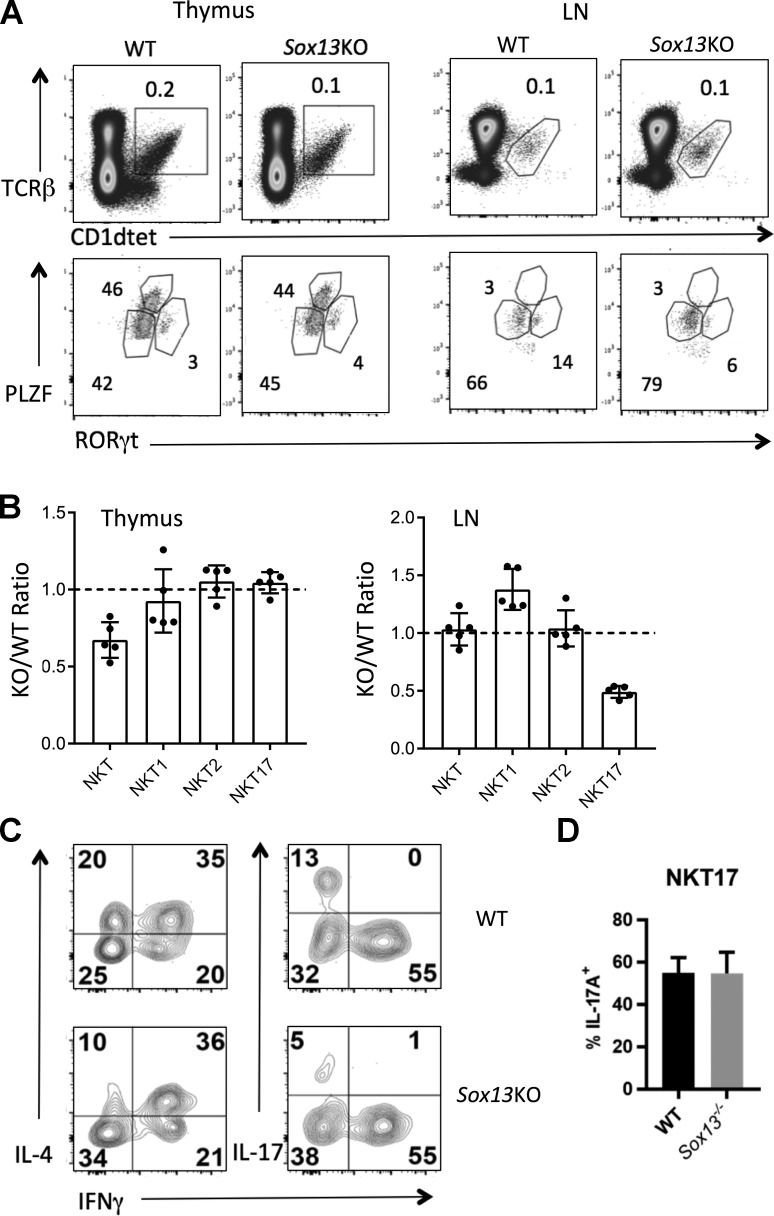
Natural killer T (NKT) cells expressing the invariant T cell receptor (iTCR) serve an essential function in clearance of certain pathogens and have been implicated in autoimmune and allergic diseases. Complex effector programs of these iNKT cells are wired in the thymus, and upon thymic egress, they can respond within hours of antigenic challenges, classifying iNKT cells as innate-like. It has been assumed that the successful rearrangement of the invariant iTCRα chain is the central event in the divergence of immature thymocytes to the NKT cell lineage, but molecular properties that render the iTCR signaling distinct to permit the T cell lineage diversification remain obscure. Here we show that the High Mobility Group (HMG) transcription factor (TF) SOX4 controls the production of iNKT cells by inducing MicroRNA-181 (Mir181) to enhance TCR signaling and Ca2+ fluxes in precursors. These results suggest the existence of tailored, permissive gene circuits in iNKT precursors for innate-like T cell development.
© 2018 Malhotra et al.
Figures
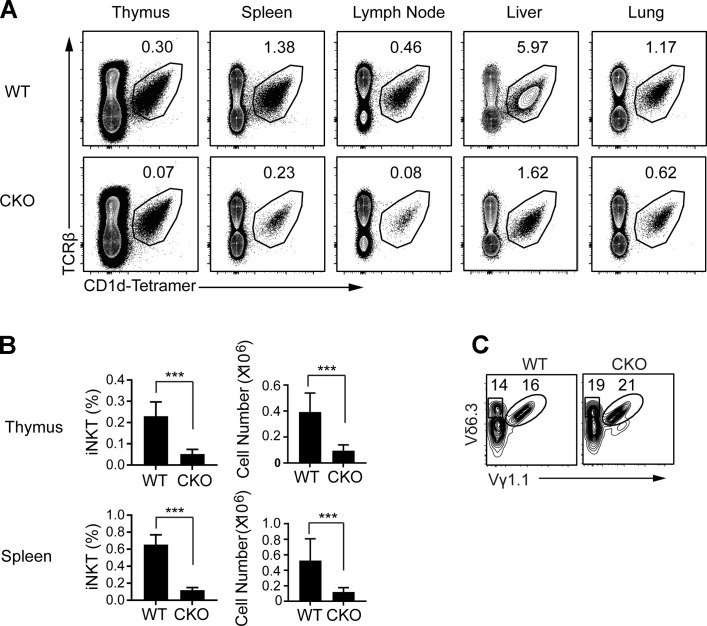
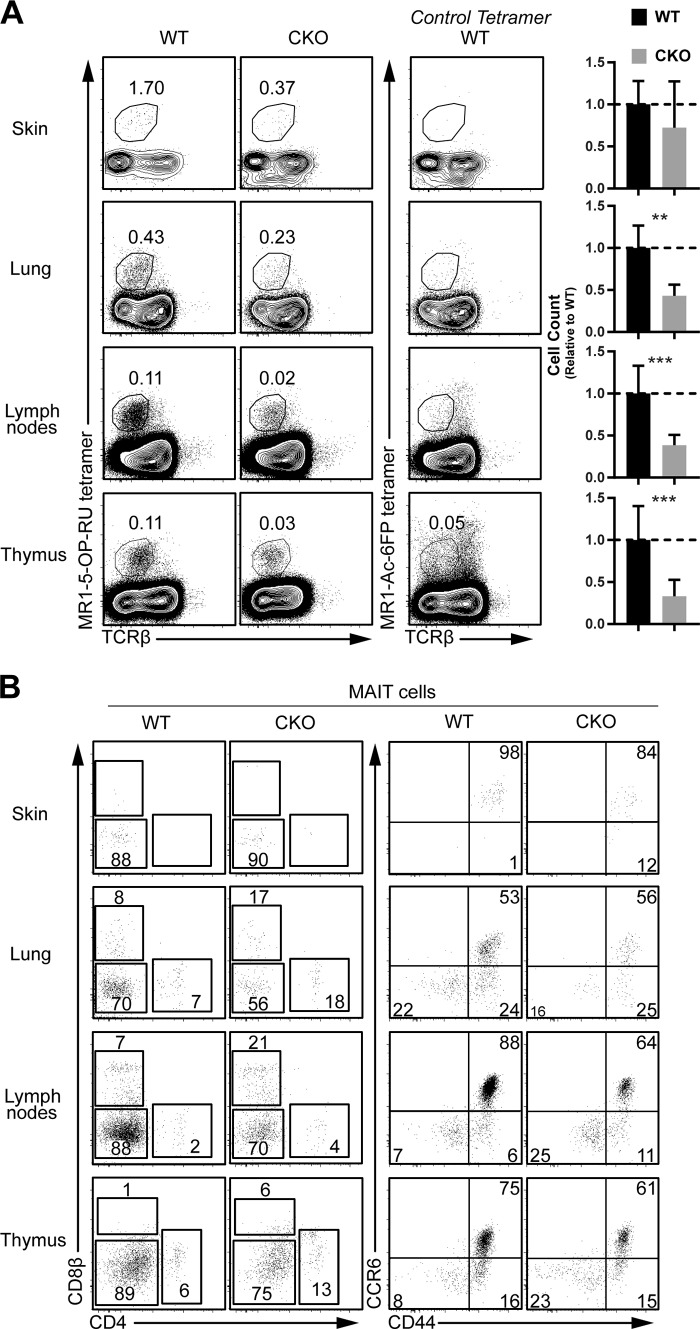
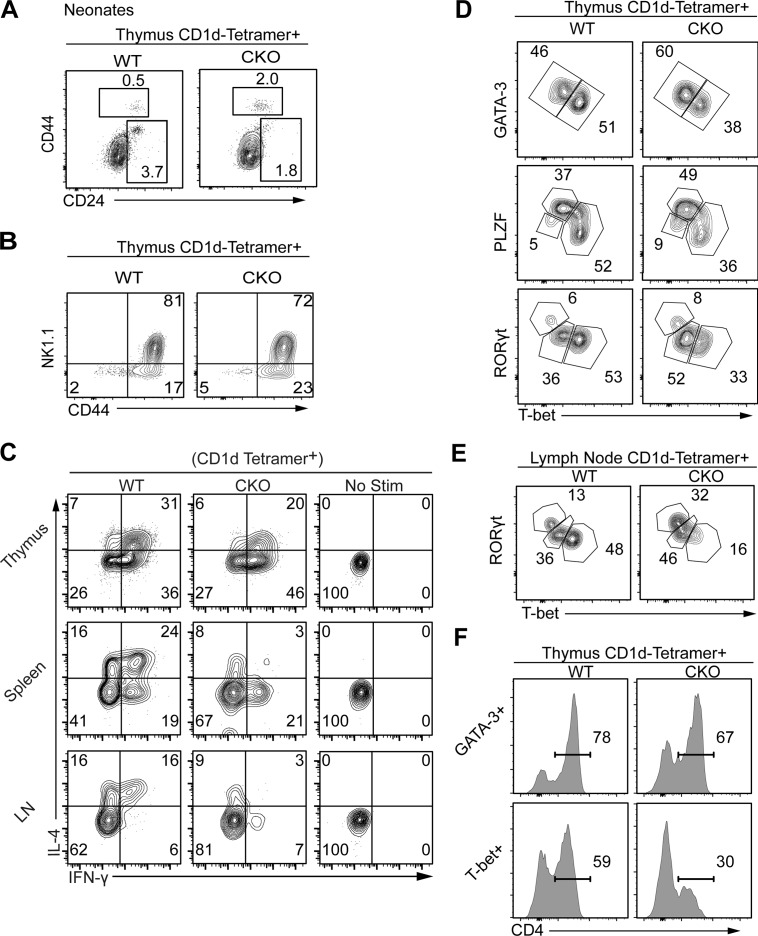
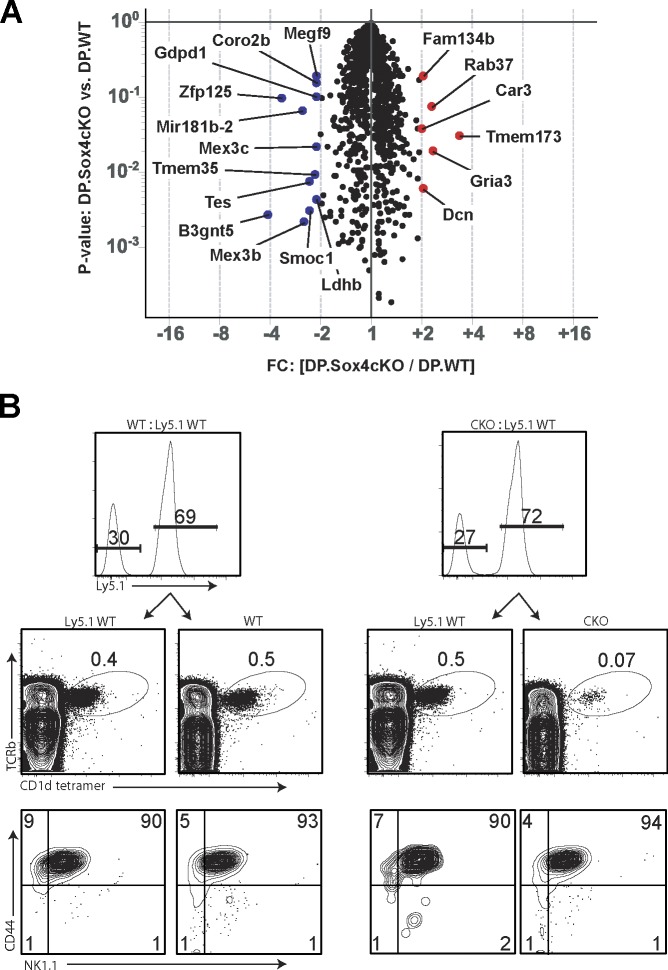

References
-
- Arase H., Ono S., Arase N., Park S.Y., Wakizaka K., Watanabe H., Ohno H., and Saito T.. 1995. Developmental arrest of NK1.1+ T cell antigen receptor (TCR)-alpha/beta+ T cells and expansion of NK1.1+ TCR-gamma/delta+ T cell development in CD3 zeta-deficient mice. J. Exp. Med. 182:891–895. 10.1084/jem.182.3.891 - DOI - PMC - PubMed
-
- Carnaud C., Lee D., Donnars O., Park S.-H., Beavis A., Koezuka Y., and Bendelac A.. 1999. Cutting edge: Cross-talk between cells of the innate immune system: NKT cells rapidly activate NK cells. J. Immunol. 163:4647–4650. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
