

A point mutation in the Ncr1 signal peptide impairs the development of innate lymphoid cell subsets
- PMID: 30288342
- PMCID: PMC6169588
- DOI: 10.1080/2162402X.2018.1475875
A point mutation in the Ncr1 signal peptide impairs the development of innate lymphoid cell subsets
Abstract
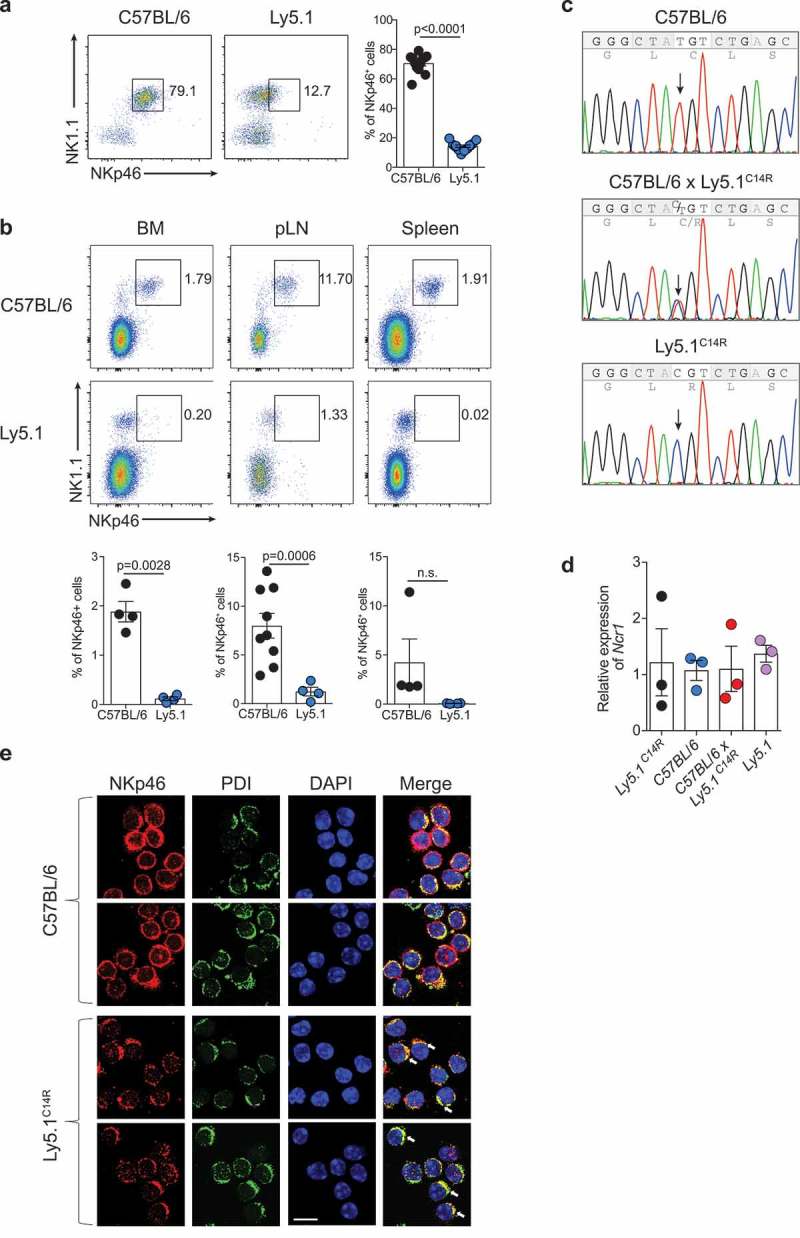
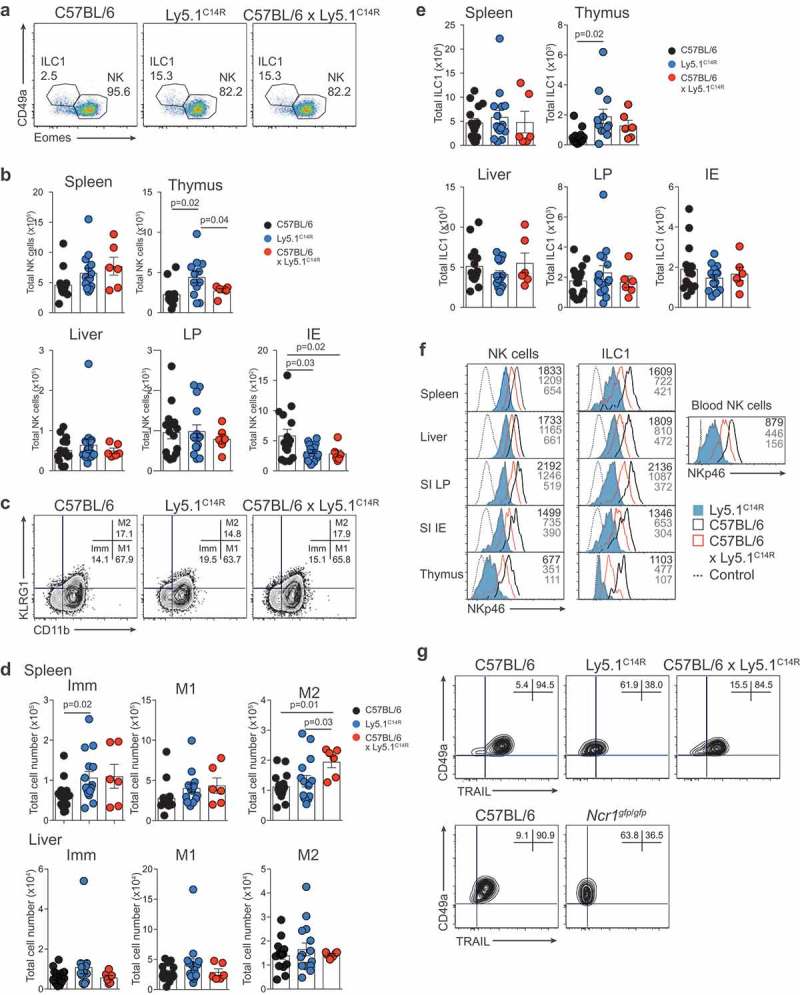
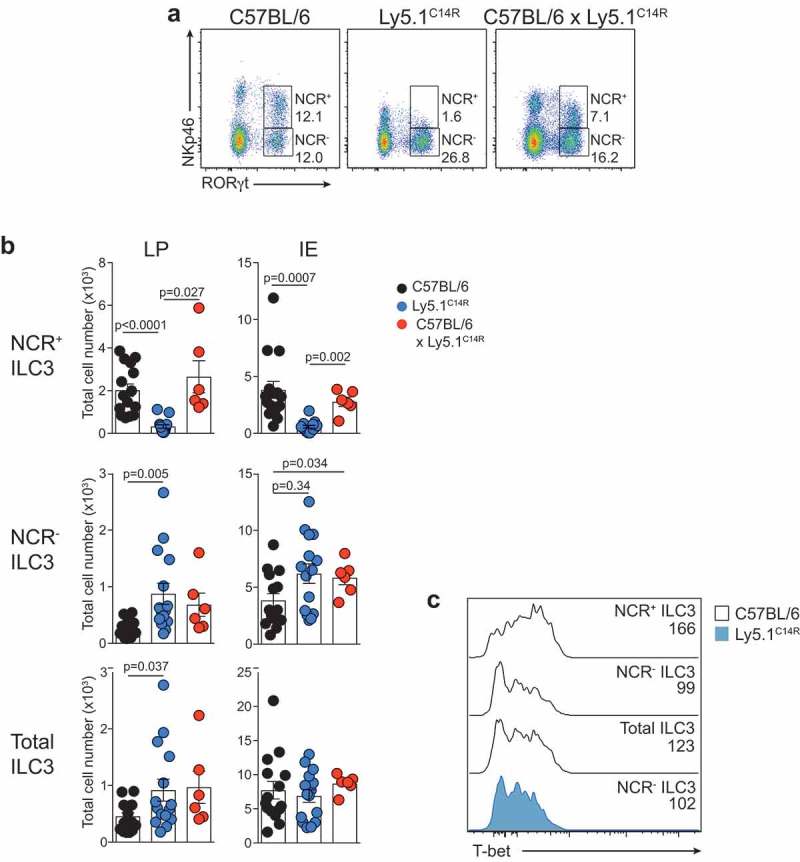
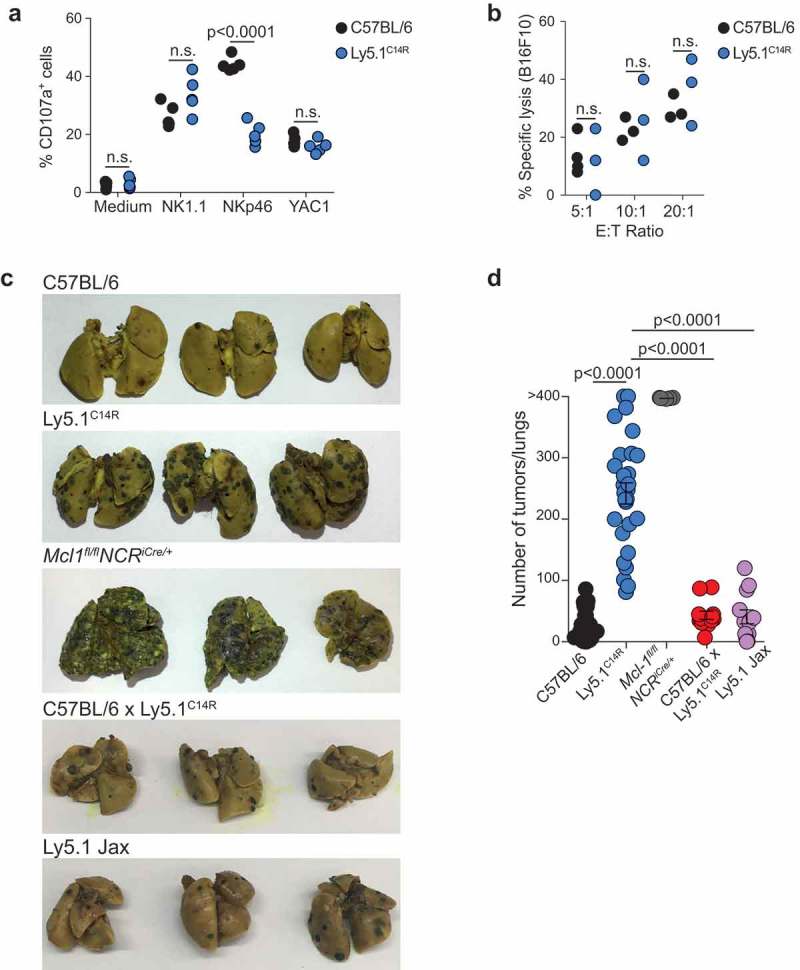
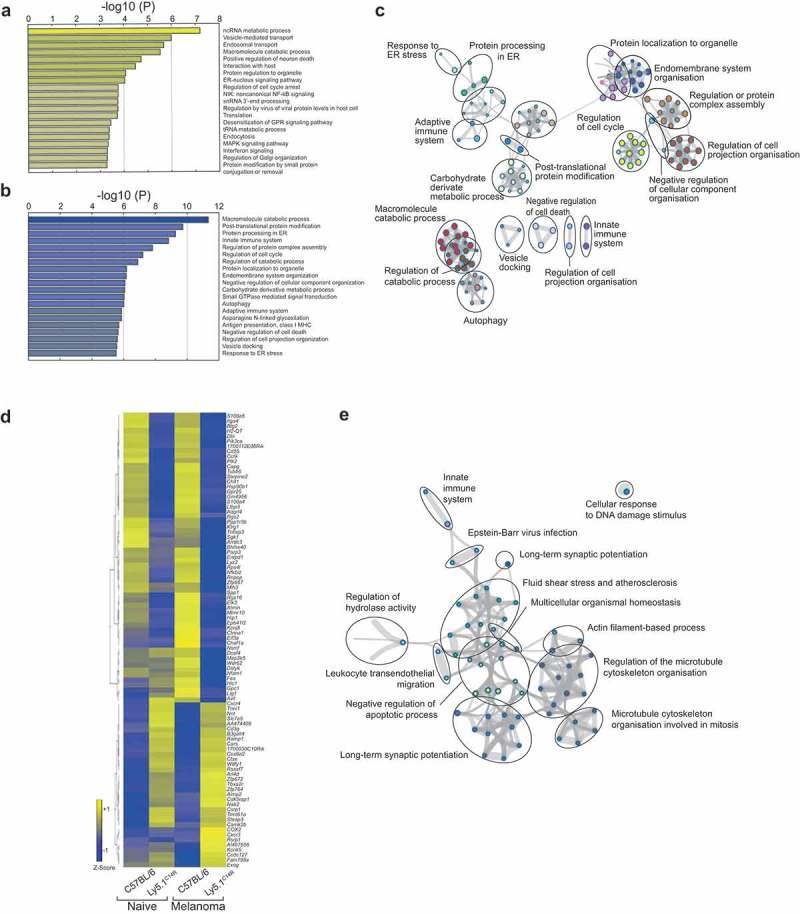
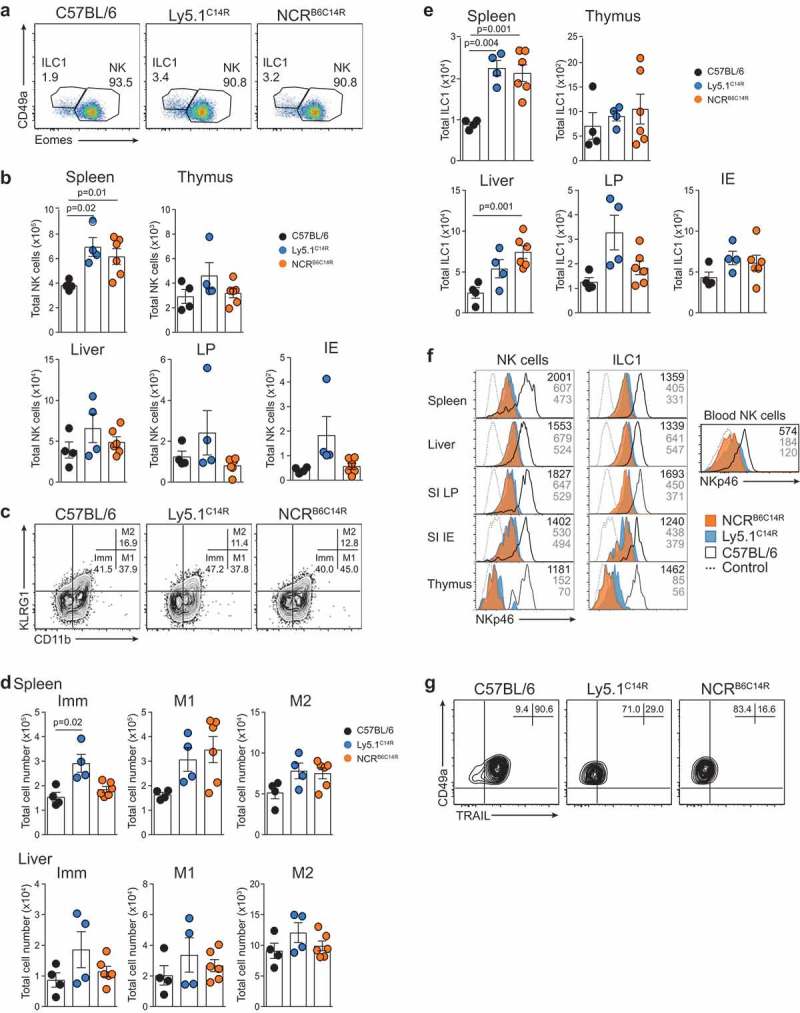
NKp46 (CD335) is a surface receptor shared by both human and mouse natural killer (NK) cells and innate lymphoid cells (ILCs) that transduces activating signals necessary to eliminate virus-infected cells and tumors. Here, we describe a spontaneous point mutation of cysteine to arginine (C14R) in the signal peptide of the NKp46 protein in congenic Ly5.1 mice and the newly generated NCRB6C14R strain. Ly5.1C14R NK cells expressed similar levels of Ncr1 mRNA as C57BL/6, but showed impaired surface NKp46 and reduced ability to control melanoma tumors in vivo. Expression of the mutant NKp46C14R in 293T cells showed that NKp46 protein trafficking to the cell surface was compromised. Although Ly5.1C14R mice had normal number of NK cells, they showed an increased number of early maturation stage NK cells. CD49a+ILC1s were also increased but these cells lacked the expression of TRAIL. ILC3s that expressed NKp46 were not detectable and were not apparent when examined by T-bet expression. Thus, the C14R mutation reveals that NKp46 is important for NK cell and ILC differentiation, maturation and function. Significance Innate lymphoid cells (ILCs) play important roles in immune protection. Various subsets of ILCs express the activating receptor NKp46 which is capable of recognizing pathogen derived and tumor ligands and is necessary for immune protection. Here, we describe a spontaneous point mutation in the signal peptide of the NKp46 protein in congenic Ly5.1 mice which are widely used for tracking cells in vivo. This Ncr1 C14R mutation impairs NKp46 surface expression resulting in destabilization of Ncr1 and accumulation of NKp46 in the endoplasmic reticulum. Loss of stable NKp46 expression impaired the maturation of NKp46+ ILCs and altered the expression of TRAIL and T-bet in ILC1 and ILC3, respectively.
Keywords: activation receptors; congenic mice; innate lymphoid cells; intracellular trafficking.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials