

HMGA2 as a functional antagonist of PARP1 inhibitors in tumor cells
- PMID: 30289618
- PMCID: PMC6360374
- DOI: 10.1002/1878-0261.12390
HMGA2 as a functional antagonist of PARP1 inhibitors in tumor cells
Abstract
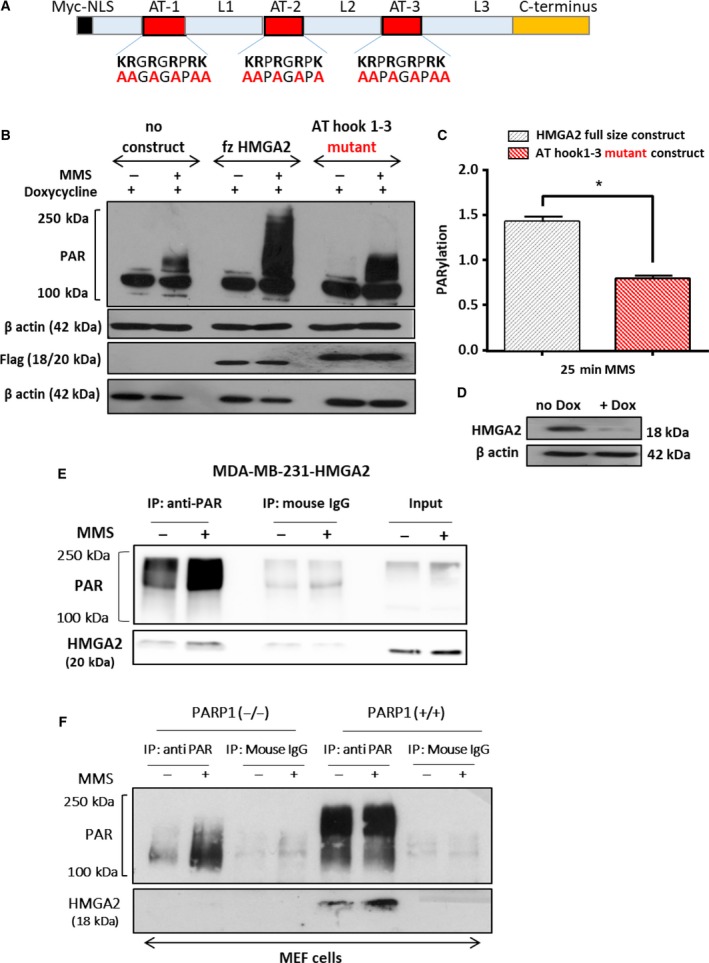
Poly(ADP-ribose) polymerase 1 inhibitors alone or in combination with DNA damaging agents are promising clinical drugs in the treatment of cancer. However, there is a need to understand the molecular mechanisms of resistance to PARP1 inhibitors. Expression of HMGA2 in cancer is associated with poor prognosis for patients. Here, we investigated the novel relationship between HMGA2 and PARP1 in DNA damage-induced PARP1 activity. We used human triple-negative breast cancer and fibrosarcoma cell lines to demonstrate that HMGA2 colocalizes and interacts with PARP1. High cellular HMGA2 levels correlated with increased DNA damage-induced PARP1 activity, which was dependent on functional DNA-binding AT-hook domains of HMGA2. HMGA2 inhibited PARP1 trapping to DNA and counteracted the cytotoxic effect of PARP inhibitors. Consequently, HMGA2 decreased caspase 3/7 induction and increased cell survival upon treatment with the alkylating methyl methanesulfonate alone or in combination with the PARP inhibitor AZD2281 (olaparib). HMGA2 increased mitochondrial oxygen consumption rate and spare respiratory capacity and increased NAMPT levels, suggesting metabolic support for enhanced PARP1 activity upon DNA damage. Our data showed that expression of HMGA2 in cancer cells reduces sensitivity to PARP inhibitors and suggests that targeting HMGA2 in combination with PARP inhibition may be a promising new therapeutic approach.
Keywords: HMGA2; PARP1; PARP1 trapping; PARylation; olaparib.
© 2018 The Authors. Published by FEBS Press and John Wiley & Sons Ltd.
Figures





References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
