

Mutations in KCNK4 that Affect Gating Cause a Recognizable Neurodevelopmental Syndrome
- PMID: 30290154
- PMCID: PMC6174320
- DOI: 10.1016/j.ajhg.2018.09.001
Mutations in KCNK4 that Affect Gating Cause a Recognizable Neurodevelopmental Syndrome
Abstract
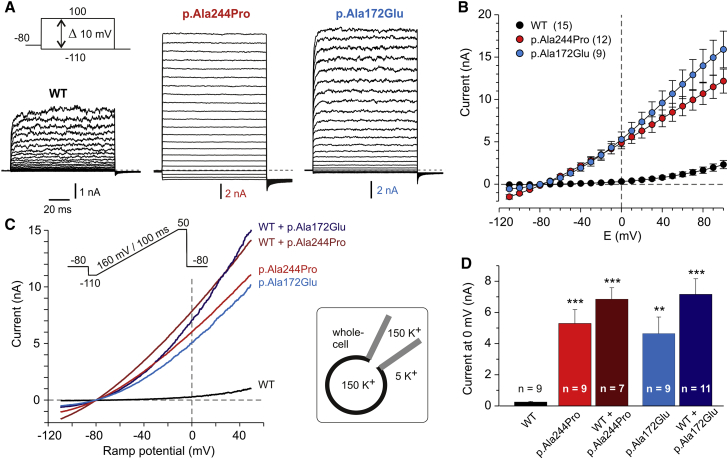
Aberrant activation or inhibition of potassium (K+) currents across the plasma membrane of cells has been causally linked to altered neurotransmission, cardiac arrhythmias, endocrine dysfunction, and (more rarely) perturbed developmental processes. The K+ channel subfamily K member 4 (KCNK4), also known as TRAAK (TWIK-related arachidonic acid-stimulated K+ channel), belongs to the mechano-gated ion channels of the TRAAK/TREK subfamily of two-pore-domain (K2P) K+ channels. While K2P channels are well known to contribute to the resting membrane potential and cellular excitability, their involvement in pathophysiological processes remains largely uncharacterized. We report that de novo missense mutations in KCNK4 cause a recognizable syndrome with a distinctive facial gestalt, for which we propose the acronym FHEIG (facial dysmorphism, hypertrichosis, epilepsy, intellectual disability/developmental delay, and gingival overgrowth). Patch-clamp analyses documented a significant gain of function of the identified KCNK4 channel mutants basally and impaired sensitivity to mechanical stimulation and arachidonic acid. Co-expression experiments indicated a dominant behavior of the disease-causing mutations. Molecular dynamics simulations consistently indicated that mutations favor sealing of the lateral intramembrane fenestration that has been proposed to negatively control K+ flow by allowing lipid access to the central cavity of the channel. Overall, our findings illustrate the pleiotropic effect of dysregulated KCNK4 function and provide support to the hypothesis of a gating mechanism based on the lateral fenestrations of K2P channels.
Keywords: FHEIG syndrome; K2P channels; TRAAK; channelopathy; epilepsy; gingival overgrowth; hypertrichosis; intellectual disability; neurodevelopmental disorder.
Copyright © 2018 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Hille B. Sinauer Associates; 2001. Ion channels of excitable membranes.
-
- Tian C., Zhu R., Zhu L., Qiu T., Cao Z., Kang T. Potassium channels: structures, diseases, and modulators. Chem. Biol. Drug Des. 2014;83:1–26. - PubMed
-
- Shieh C.C., Coghlan M., Sullivan J.P., Gopalakrishnan M. Potassium channels: molecular defects, diseases, and therapeutic opportunities. Pharmacol. Rev. 2000;52:557–594. - PubMed
-
- Jentsch T.J. Neuronal KCNQ potassium channels: physiology and role in disease. Nat. Rev. Neurosci. 2000;1:21–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
