

A dominant dendrite phenotype caused by the disease-associated G253D mutation in doublecortin (DCX) is not due to its endocytosis defect
- PMID: 30291144
- PMCID: PMC6295730
- DOI: 10.1074/jbc.RA118.004462
A dominant dendrite phenotype caused by the disease-associated G253D mutation in doublecortin (DCX) is not due to its endocytosis defect
Abstract
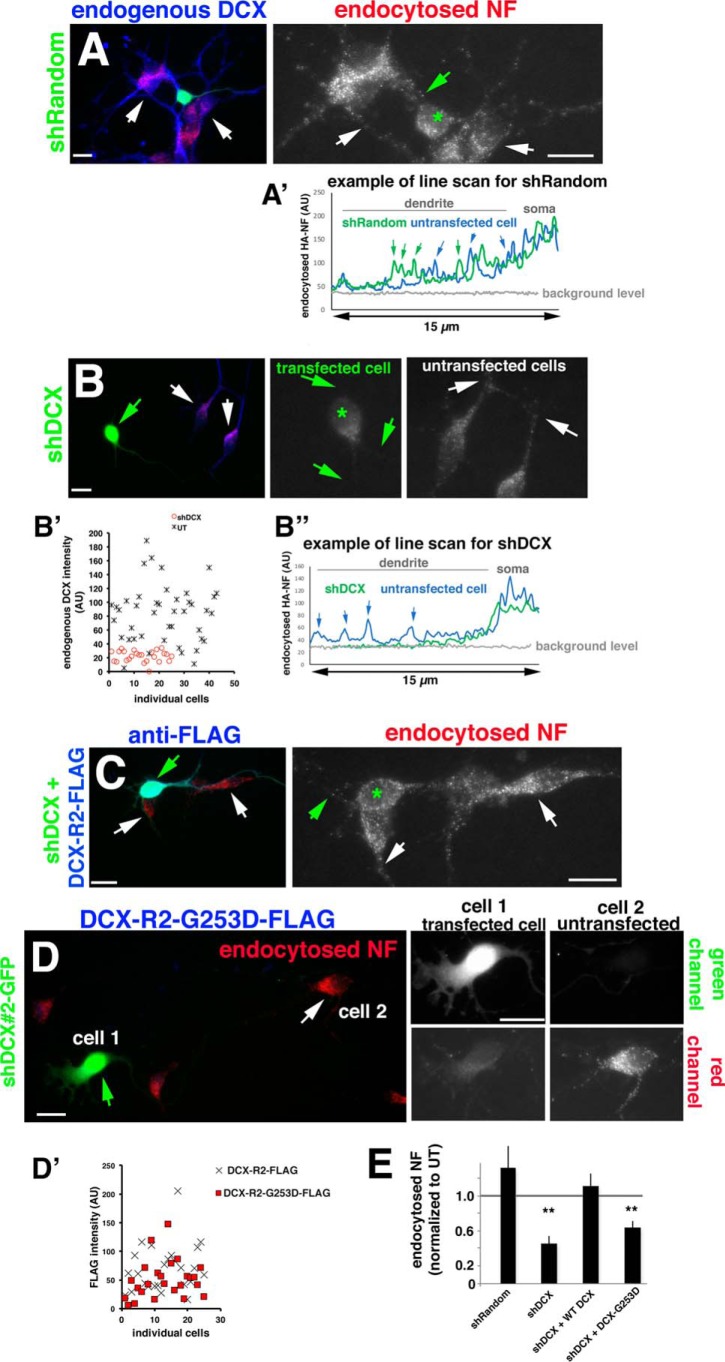
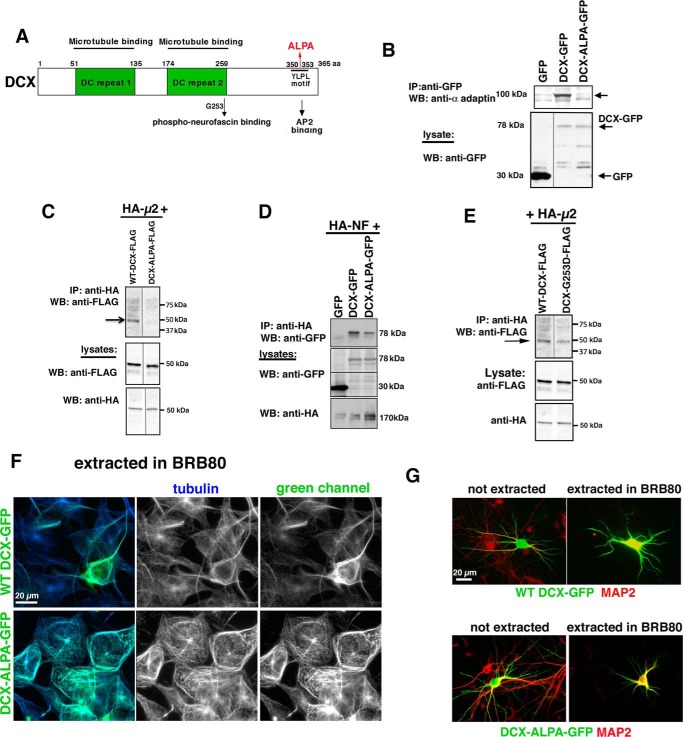
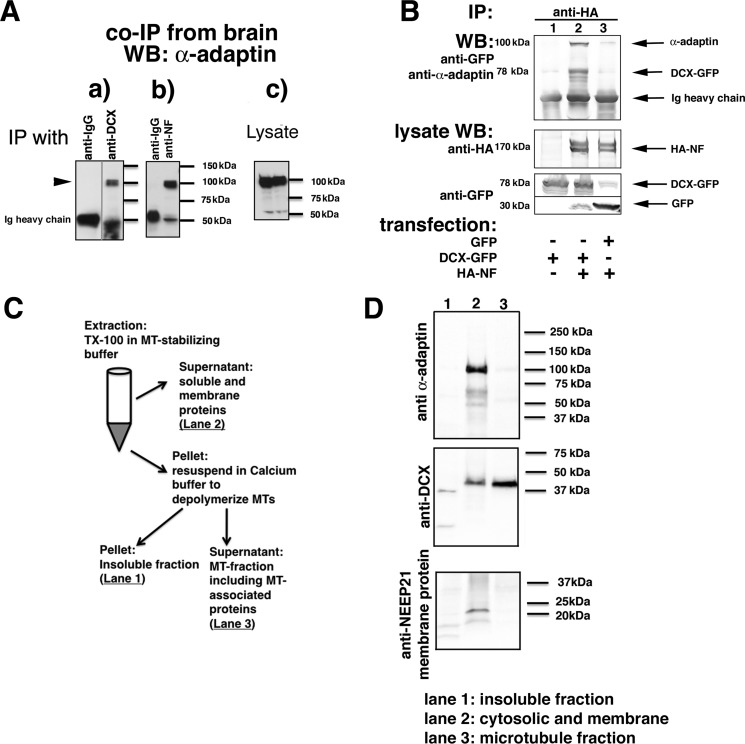
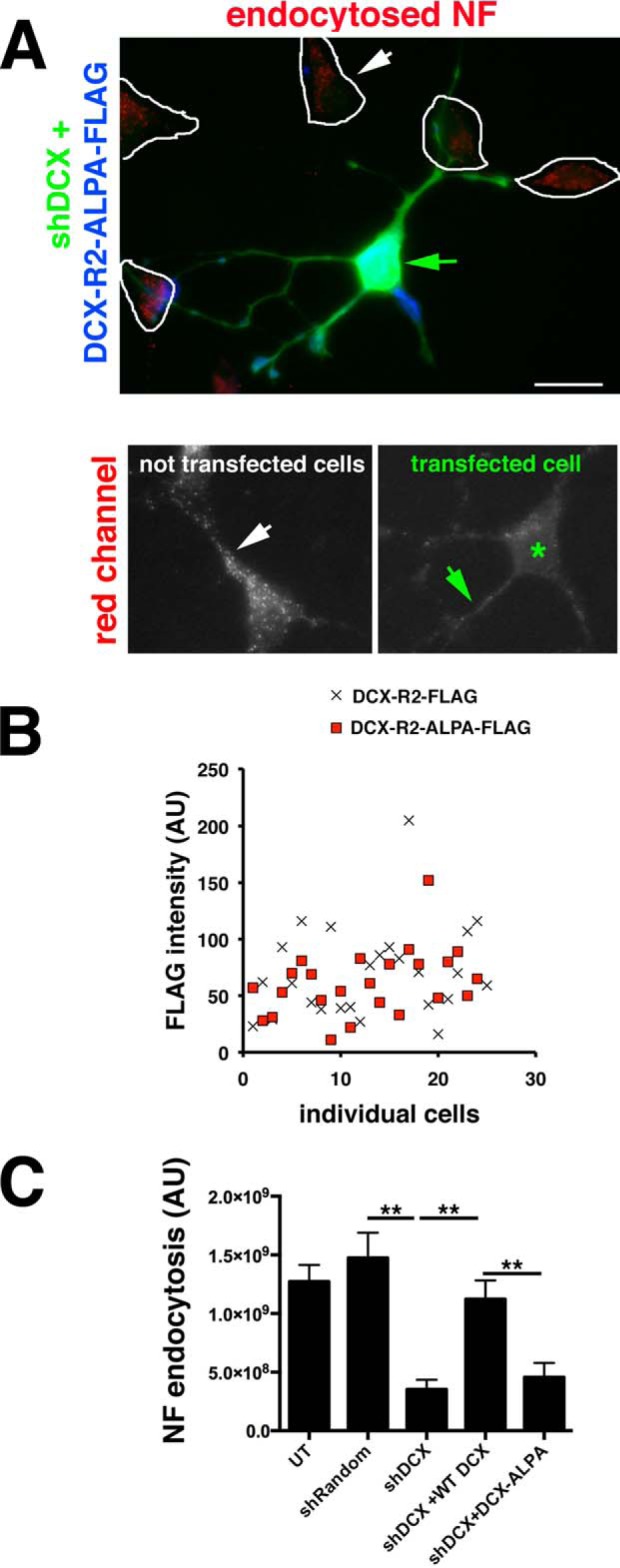
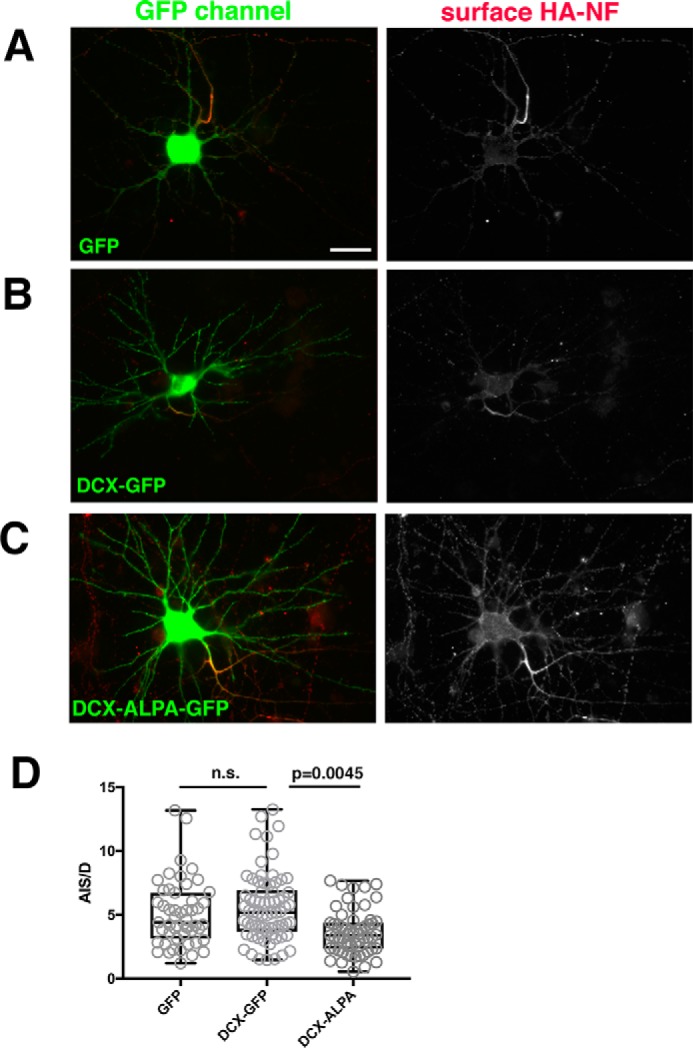
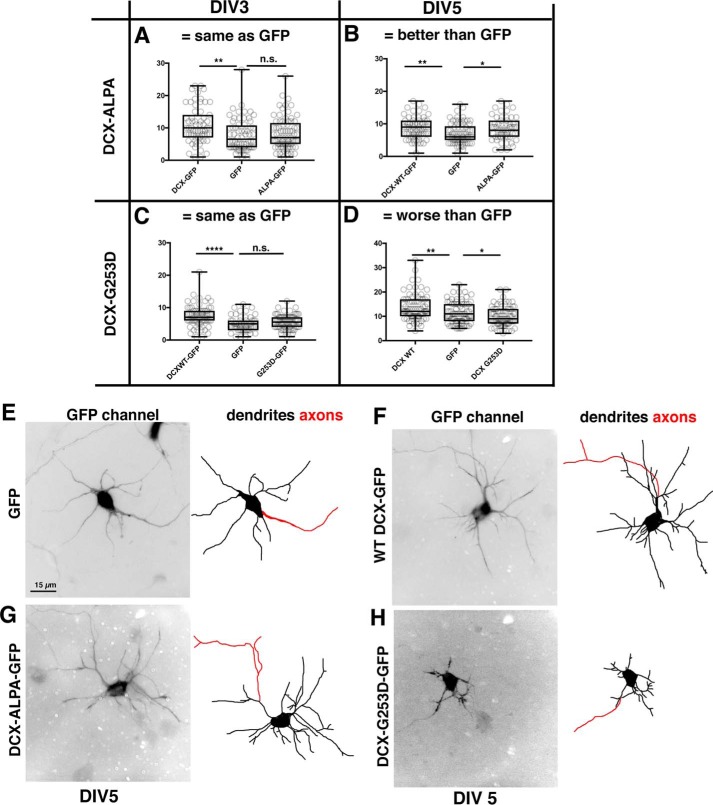
Doublecortin (DCX) is a protein needed for cortical development, and DCX mutations cause cortical malformations in humans. The microtubule-binding activity of DCX is well-described and is important for its function, such as supporting neuronal migration and dendrite growth during development. Previous work showed that microtubule binding is not sufficient for DCX-mediated promotion of dendrite growth and that domains in DCX's C terminus are also required. The more C-terminal regions of DCX bind several other proteins, including the adhesion receptor neurofascin and clathrin adaptors. We recently identified a role for DCX in endocytosis of neurofascin. The disease-associated DCX-G253D mutant protein is known to be deficient in binding neurofascin, and we now asked if disruption of neurofascin endocytosis underlies the DCX-G253D-associated pathology. We first demonstrated that DCX functions in endocytosis as a complex with both the clathrin adaptor AP-2 and neurofascin: disrupting either clathrin adaptor binding (DCX-ALPA) or neurofascin binding (DCX-G253D) decreased neurofascin endocytosis in primary neurons. We then investigated a known function for DCX, namely, increasing dendrite growth in cultured neurons. Surprisingly, we found that the DCX-ALPA and DCX-G253D mutants yield distinct dendrite phenotypes. Unlike DCX-ALPA, DCX-G253D caused a dominant-negative dendrite growth phenotype. The endocytosis defect of DCX-G253D thus was separable from its detrimental effects on dendrite growth. We recently identified Dcx-R59H as a dominant allele and can now classify Dcx-G253D as a second Dcx allele that acts dominantly to cause pathology, but does so via a different mechanism.
Keywords: axon initial segment; clathrin; clathrin adaptor; dendrite; dominant allele of DCX; endocytosis; lissencephaly; microtubule; multifunctional protein; neurite outgrowth; neurofascin; neuron.
© 2018 Yap et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
