

Seasonal changes in neuronal turnover in a forebrain nucleus in adult songbirds
- PMID: 30291632
- PMCID: PMC6333494
- DOI: 10.1002/cne.24552
Seasonal changes in neuronal turnover in a forebrain nucleus in adult songbirds
Abstract
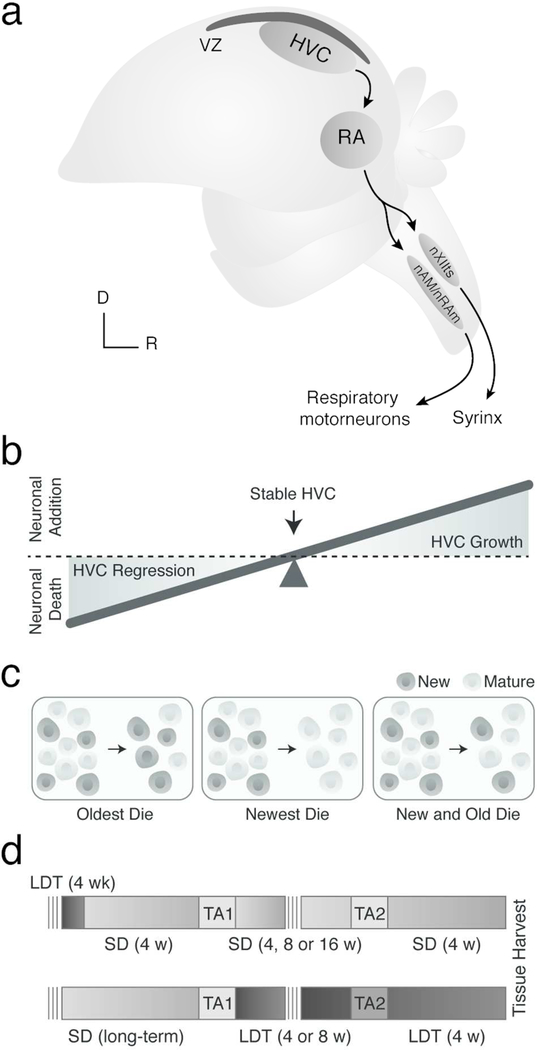
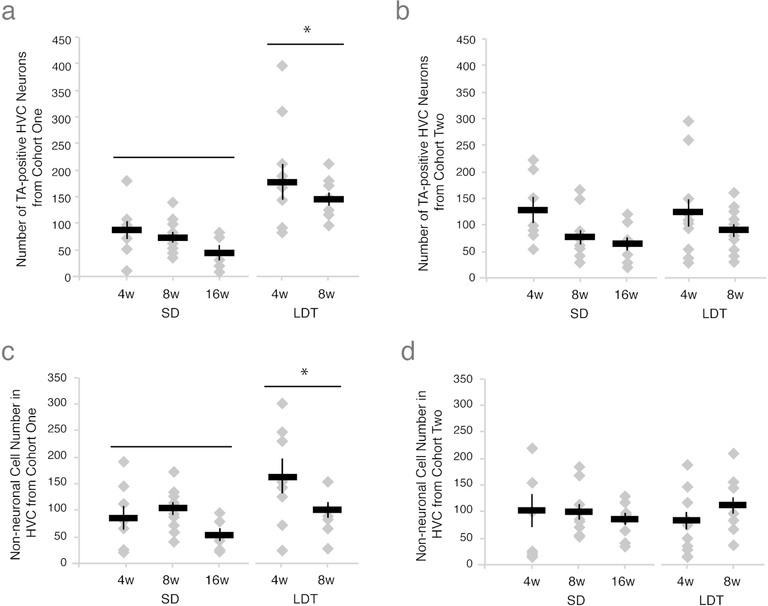
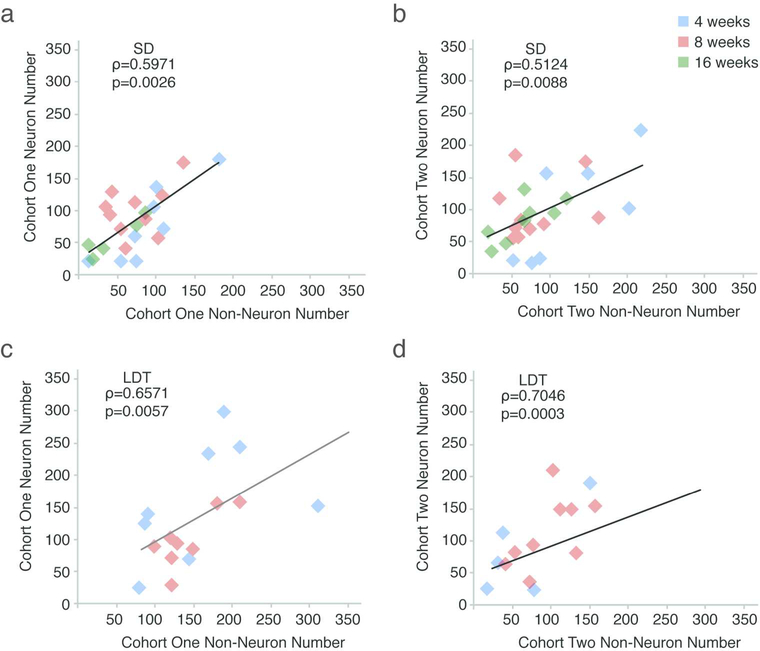
Neuronal death and replacement, or neuronal turnover, in the adult brain are one of many fundamental processes of neural plasticity. The adult avian song control circuit provides an excellent model for exploring mature neuronal death and replacement by new neurons. In the song control nucleus, HVC of adult male Gambel's white-crowned sparrows (Zonotrichia leucophrys gambelli) nearly 68,000 neurons are added each breeding season and die during the subsequent nonbreeding season. To accommodate large seasonal differences in HVC neuron number, the balance between neuronal addition and death in HVC must differ between seasons. To determine whether maintenance of new HVC neurons changes within and between breeding and nonbreeding conditions, we pulse-labeled two different cohorts of new HVC neurons under both conditions and quantified their maintenance. We show that the maintenance of new HVC neurons, as well as new nonneuronal cells, was higher at the onset of breeding conditions than at the onset of nonbreeding conditions. Once a steady-state HVC volume and neuronal number were attained in either breeding or nonbreeding conditions, neuronal and nonneuronal maintenance were similarly low. We found that new neuronal number correlated with a new nonneuronal number within each cohort of new neurons. Together, these data suggest that sex steroids promote the survival of an initial population of new neurons and nonneuronal cells entering HVC. However, once HVC is fully grown or regressed, neuronal and nonneuronal cell turnover is regulated by a common mechanism likely independent of direct sex steroid signaling.
Keywords: adult neurogenesis; anti-BrdU (RRID: AB_2536432); anti-NeuN (RRID: AB_2532109); degeneration; neuronal death; seasonal plasticity; songbird; steroid hormone; testosterone.
© 2018 Wiley Periodicals, Inc.
Conflict of interest statement
Figures
References
-
- Alvarez-Buylla A, Ling CY, Yu WS. 1994. Contribution of neurons born during embryonic, juvenile, and adult life to the brain of adult canaries: regional specificity and delayed birth of neurons in the song-control nuclei. J Comp Neurol 347(2):233–248. - PubMed
-
- Alvarez-Buylla A, Theelen M, Nottebohm F. 1990. Proliferation “hot spots” in adult avian ventricular zone reveal radial cell division. Neuron 5(1):101–109. - PubMed
-
- Bentley GE, Audage NC, Hanspal EK, Ball GF, Hahn TP. 2003. Photoperiodic response of the hypothalamo-pituitary-gonad axis in male and female canaries, Serinus canaria. J Exp Zool A Comp Exp Biol 296(2):143–151. - PubMed
-
- Brenowitz EA. 2008. Plasticity of the song control system in adult birds In: Zeigler HP, Marler P, eds. Neuroscience of birdsong. Cambridge: Cambridge University Press; p 332–349.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
