

Review
. 2018 Oct;6(5):10.1128/microbiolspec.gpp3-0026-2018.
doi: 10.1128/microbiolspec.GPP3-0026-2018.
Mycobacteriophages
Affiliations
- PMID: 30291704
- PMCID: PMC6282025
- DOI: 10.1128/microbiolspec.GPP3-0026-2018
Item in Clipboard
Review
Mycobacteriophages
Microbiol Spectr.
2018 Oct.
Abstract
Mycobacteriophages are viruses that infect mycobacterial hosts. A large number of mycobacteriophages have been isolated and genomically characterized, providing insights into viral diversity and evolution, as well as fueling development of tools for mycobacterial genetics. Mycobacteriophages have intimate relationships with their hosts and provide insights into the genetics and physiology of the mycobacteria and tools for potential clinical applications such as drug development, diagnosis, vaccines, and potentially therapy.
Figures

Organization and structure of the SEA-PHAGES program. SEA-PHAGES program administrators (yellow box, top) oversee support components critical to program implementation (green box, upper middle). The typical two-term course structure (red box, lower middle) includes phage isolation through comparative genomics; additional characterization includes electron microscopy, PCR/restriction analysis, and lysogeny assays (red ovals). Sequence and annotation quality control is shared with SEA-PHAGES faculty teams (purple box, bottom right). Reproduced with permission from reference .

Mycobacteriophage viral morphotypes. Mycobacteriophages generally fall into two morphological families, the Siphoviridae with long flexible tails (e.g., Phaedrus and Catdawg) and the Myoviridae with contractile tails (e.g., Rizal). Most of the siphoviral phages have isometric heads (e.g., Phaedrus), but some (e.g., Catdawg) have prolate heads. Scale marker is 100 nm.

Genome organization of mycobacteriophage Taheera. The genome of phage Taheera (cluster G) is represented as a scale bar (major intervals: 1 kbp) with predicted genes shown as boxes either above (rightward transcribed) or below (leftward transcribed). Gene numbers are shown within each box, and the phamily designation is shown either above or below with the number of phamily members shown in parentheses. Putative gene functions are indicated.

Gene expression in mycobacteriophage Fruitloop. RNA was isolated from M. smegmatis infected with phage Fruitloop at either early (30 min) or late (150 min) time points after infection. Following strand-specific RNAseq analysis, sequence reads were mapped to the Fruitloop genome (shown below). Reads mapping to the forward and reverse strands are indicated on the right. Scales for the numbers of sequence reads are from 0 to 5,000. Reproduced with permission from reference .

Integration-dependent immunity regulation. BPs is a cluster G phage, and a central ∼4 kbp segment of the genome is shown at the top. Genes are shown as colored boxes, transcribed rightward or leftward as depicted above or below the genome, respectively. The repressor and integrase genes (33 and 32, respectively) are transcribed leftward from the Prep promoter. The attP site (black bar) is located within the repressor open reading frame and can recombine with an attB site overlapping an M. smegmatis tRNAarg gene. Establishment of lysogeny involves formation of an integrated prophage in which an active form of the repressor is expressed from near attL, and the 3′ end of the gene is near attR. The virally encoded form of the repressor contains a C-terminal ssrA-like tag that targets it for degradation, and it fails to confer immunity. Integration results in removal of the C-terminal tag such that the prophage-expressed form of the repressor is stable and confers immunity.

Gene expression in a Panchino lysogen of M. smegmatis. RNA was isolated from a lysogen of mycobacteriophage Panchino and strand-specific RNAseq reads were mapped to the Panchino prophage. Only the genome segments at the ends of the prophage near attL and attR are shown. Sequence reads for forward and reverse DNA direction are shown, and the scales are 0 to 500 reads. Genes below the genome are transcribed leftward, and those above are transcribed rightward. Panchino uses an integration-dependent immunity system similar to that in phage BPs (see Fig. 5). Genes 31 to 28 are also expressed; the lytic genes are not expressed (not shown). Reproduced with permission from reference .

Genomic mosaicism in subcluster A2 phages. Mycobacteriophages are characteristically mosaic, with shared genome segments interspersed with nonhomologous regions. The genomes of 12 subcluster A2 phages are shown (Bactobuster, Che12, D29, Echild, Jaan, L5, Pukovnik, RedRock, Serenity, StarStuff, Turbido, and Updawg), with pairwise nucleotide sequence similarities shown as shadings between the genomes; shading is spectrum colored, with violet being the most closely related and red being just above the threshold E value of 10−5. Genes are shown as colored boxes, and genes within the same phamily are similarly colored. Note the segments with close sequence similarity (violet-shaded regions) interspersed with dissimilar regions (white-shaded segments).

Single-gene mosaics in mycobacteriophage genomes. Segments of five phage genomes (Phrann, Corndog, Brujita, Squirty, and Gaia) are shown, which are unrelated at the nucleotide sequence level and are grouped in different clusters, as shown (in clusters, N, O, I1, F3, and X, respectively). Genes are shown as colored boxes, with genes in the same phamilies colored similarly and pham numbers shown above with the number of phamily members in parentheses. Pairwise-shared homologues are indicated with dumbbells. Note that the homologues are situated in different genomic contexts and the flanking genes are not homologous.

Modes of phage evolution. (A) Nucleotide distance (using Mash) and gene content dissimilarity (using phams from Phamerator) are plotted for ∼2.4 × 106 dsDNA phage comparisons to reveal two evolutionary modes, HGCF and LGCF (inset). The line at y = 2x is plotted for reference. Marginal frequency histograms emphasize densely plotted regions, with truncated y axes for viewability. (B–D) Cluster-specific intracluster (orange) and intercluster (black) comparisons are plotted as in Fig. 1A for actinobacteriophage clusters B (panel B), F (panel C), and K (panel D). Phages in clusters F and K are temperate, and phages in cluster B are lytic, as indicated. n, number of phages present in the specific cluster. Reproduced with permission from reference .

A model for mycobacteriophage diversity. The diversity of mycobacteriophages isolated on M. smegmatis mc2155 can be explained by a model in which (i) phages can readily infect a new bacterial host, by either a switch or an expansion of host range, and (ii) there is a highly diverse bacterial population, including many closely related strains, in the environments from which the phages are isolated. As such, phages with distinctly different genome sequences and GC contents infecting distantly related bacterial hosts, such as those to the left (red) or right (blue) extremes of a spectrum of hosts, can migrate across a microbial landscape through multiple steps. Each host switch occurs at a relatively high frequency (∼1 in 105 particles, or an average of about one every 103 bursts of lytic growth) and much faster than either amelioration of phage GC content to its new host or genetic recombination. Two phages (such as those shown in red and blue) can thus “arrive” at a common host (M. smegmatis mc2155) but be of distinctly different types (clusters, subclusters, and singletons). The variety of hosts is shown two-dimensionally for simplicity; the actual relationships among bacteria in environments such as soil and compost is likely to be considerably more complicated. Because host range variation is a common feature of bacteriophages, the model predicts that a high degree of phage diversity will be seen for any particular host if the microbial population from which the phages are isolated is also highly diverse and rich in closely related strains. Reproduced with permission from reference .

Prophage-mediated defense against phage attack. (A) Phage defense plaque assays. Ten-fold serial dilutions of phages Giles (control), MichelleMyBell (MMB), Phrann, and Tweety were plated onto lawns of a nonlysogen (mc2155) and M. smegmatis lysogens of Phrann [mc2155(Phrann)] and MichelleMyBell [mc2155(MMB)]. The Phrann and MMB lysogens confer superinfection immunity to Phrann and MMB, because these are part of cluster N, and all of the cluster N phages are homoimmune, coding for closely related repressors. Tweety (cluster F) is unrelated to Phrann and MMB, and the Phrann and MMB lysogens defend against Tweety, and the efficiency of plating is greatly reduced relative to the wild-type strain. A defense escape mutant (DEM205) overcomes the Phrann defense and plates efficiently on the Phrann lysogen but remains subject to MMB-mediated defense. DEM205 contains a mutational change in gene 54, but it is a gain of function mutant, because a Tweety mutant in which gene 54 has been deleted (Δ54) is still targeted by the Phrann defense. (B) Relationships among phage-encoded (p)ppGpp synthetase-like proteins. Phrann gp29 is a homologue of RelA/SpoT proteins with similarity to the (p)ppGpp synthetase domain of Streptococcus equisimilis RelA, including five conserved motifs (1 to 5). The cluster F phage Squirty encodes a related protein (gp29) sharing the N-terminal 124 residues with Phrann gp29 but with divergent C-termini. MMB gp29 is not predicted to be related to RelSeq but shares its C-terminus with Squirty gp29. MMB gp30 and Squirty gp30 are closely related and are both predicted to be membrane localized. (C) Models for prophage-mediated viral defense. An integrated prophage (red line) confers defense against viral attack through numerous mechanisms, either homotypically (i.e., against the same or closely related viruses) or heterotypically (i.e., against unrelated phages). Homotypic defense includes repressor-mediated immunity (repressor, red circle) and superinfection exclusion (blue circle) against itself (red phage). Heterotypic defense includes an exclusion-like system illustrated by Charlie gp32 defense against Che9c (blue phage) and restriction against many viruses (illustrated by the green phage) by Panchino gp28. Defense is also mediated by a predicted (p)ppGpp synthetase (e.g., Phrann gp29; gold circle), which we propose is kept in an inactive form (gold circle with crossed lines) by an inhibitor (purple circle), which for Squirty gp30 is membrane localized. Lytic growth by specific phages activates the defense through early lytic protein, which is proposed to dissociate the (p)ppGpp synthetase from its inhibitor, enabling rapid accumulation of (p)ppGpp and growth arrest. Tweety encodes a counterdefense system (gp54) that may prevent activation of (p)ppGpp synthesis. Reproduced with permission from reference .

Variations in tetrapeptide repeats in Tweety gp54. Gene 54 of phage Tweety codes for a 374-residue protein in which a unique N-terminal (U-L) and C-terminal (U-R) flank a long series of tetrapeptide repeats. The number of repeats in the original sequenced genome is 48, although the derivative used for defense studies (wt-N) shown here only has 40. Each of the repeats has a sequence AAXX, where XX is either WS, GY, QS, GS, or WY, and the sequence at each repeat is shown. The organization of nine defense escape mutants (DEMs) are shown below, each of which efficiently overcomes defense mediated by the Phrann prophage. They all remain subject to defense mediated by the MichelleMyBell prophage. Six of the DEM mutants (DEM10, DEM200, DEM201, DEM202, DEM204, DEM205) have lost some of the repeats but also have altered repeat sequences, as indicated. DEM202 also has an insertion of five additional repeat copies. DEM203 has two insertions, one with two repeat units and the other with four repeat units.

Genomes with potential counterdefense systems. Tweety gene 54 is active in counterdefense against Phrann (see Figs. 11, 12), and there are homologues in many other cluster F phages. The only phage outside of cluster F with a closely related homologue is the subcluster A1 phage, Nerujay. All other cluster A phages, including DD5, lack a Tweety 54 homologue. Segments of the genome maps are aligned with pairwise nucleotide sequence similarity shown as colored shading, as described for Fig. 7. In this region, all of the DD5 and Nerujay genes are transcribed leftward, and the Tweety genes are transcribed rightward.
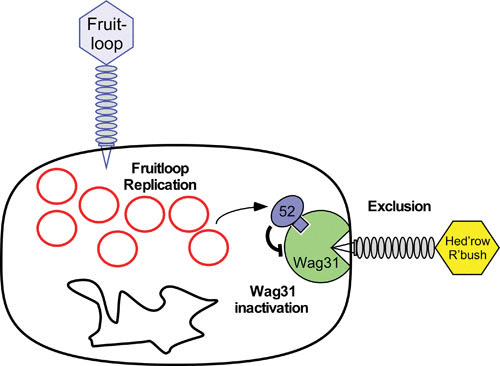
A model for Fruitloop gp52-mediated superinfection exclusion of Wag31-dependent phages. This model proposes a role for gp52 expressed by lytically growing phage Fruitloop in binding to and inactivating the host Wag31 (DivIVA) protein, which is essential and localizes to the growing pole of the cell. Some phages, such as subcluster B2 Hedgerow and Rosebush depend on Wag31 for efficient infection, such that gp52-mediated inactivation of Wag31 excludes them from superinfecting. Replicating Fruitloop genomes are shown as red circles alongside the bacterial chromosome (black circle), and Fruitloop gp52 is shown in blue, binding to Wag31, shown in green. Not drawn to scale. Reproduced with permission from reference .
References
-
- Grange JM. 1975. Proceedings: bacteriophage typing of strains of Mycobacterium tuberculosis isolated in south-east England. J Med Microbiol 8:ix. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
