

Mapping the human brain's cortical-subcortical functional network organization
- PMID: 30291974
- PMCID: PMC6289683
- DOI: 10.1016/j.neuroimage.2018.10.006
Mapping the human brain's cortical-subcortical functional network organization
Abstract
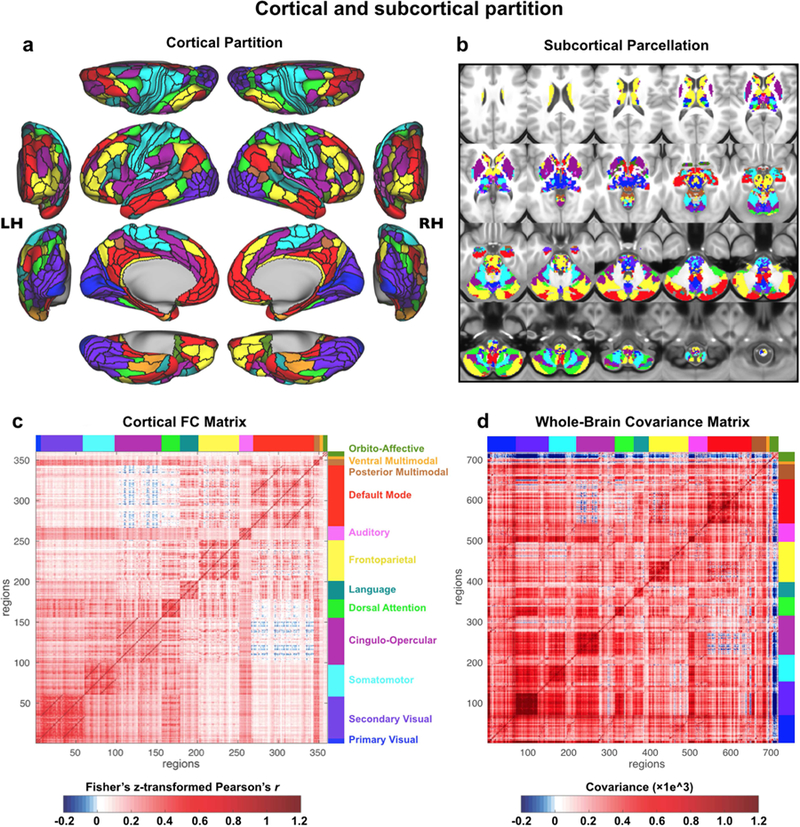
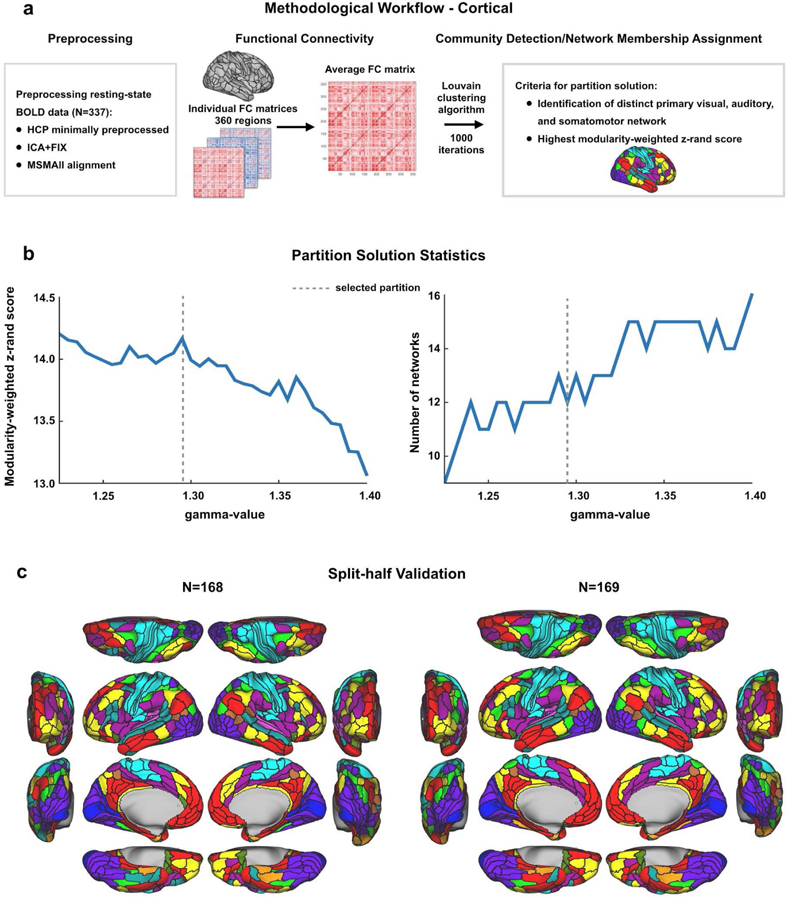
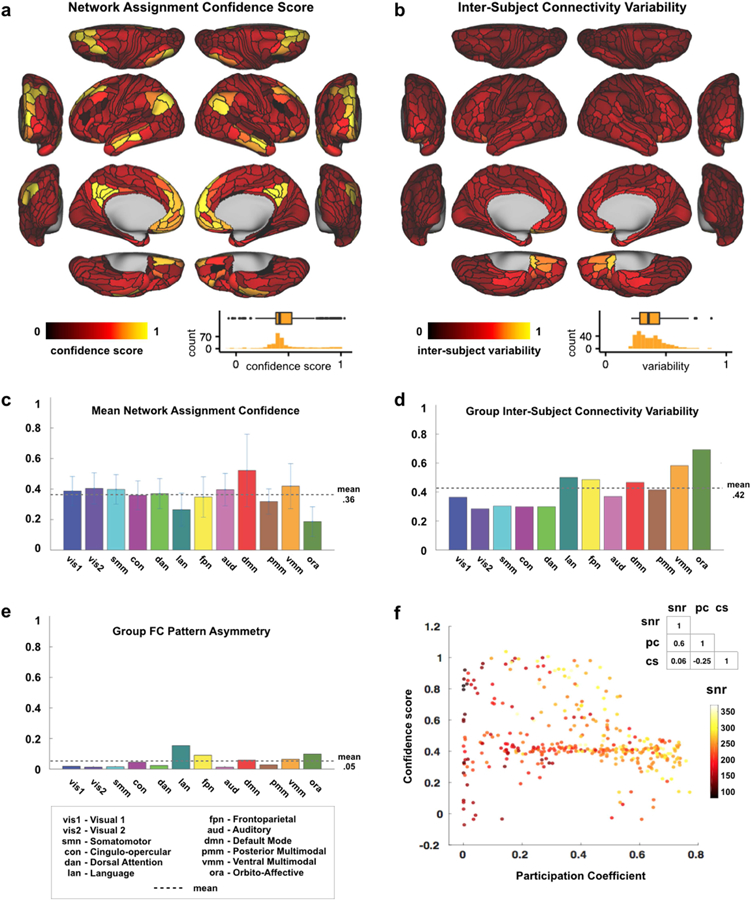
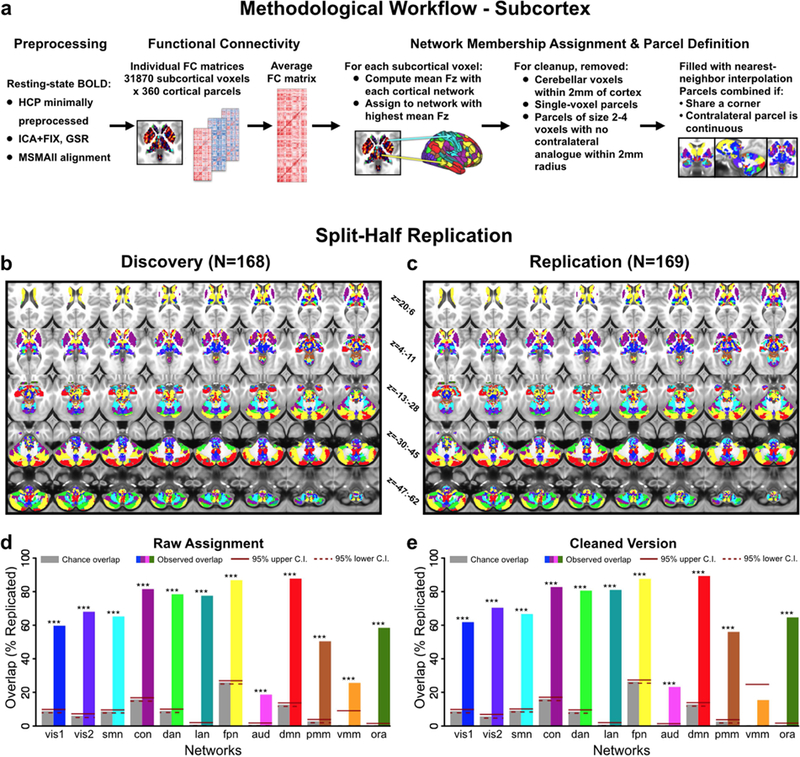
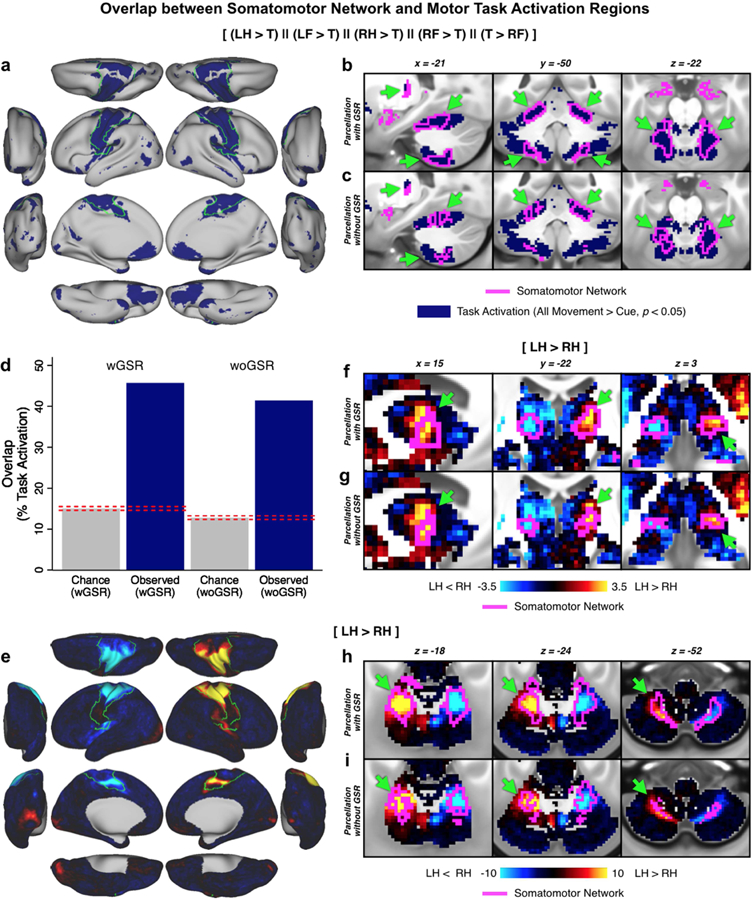
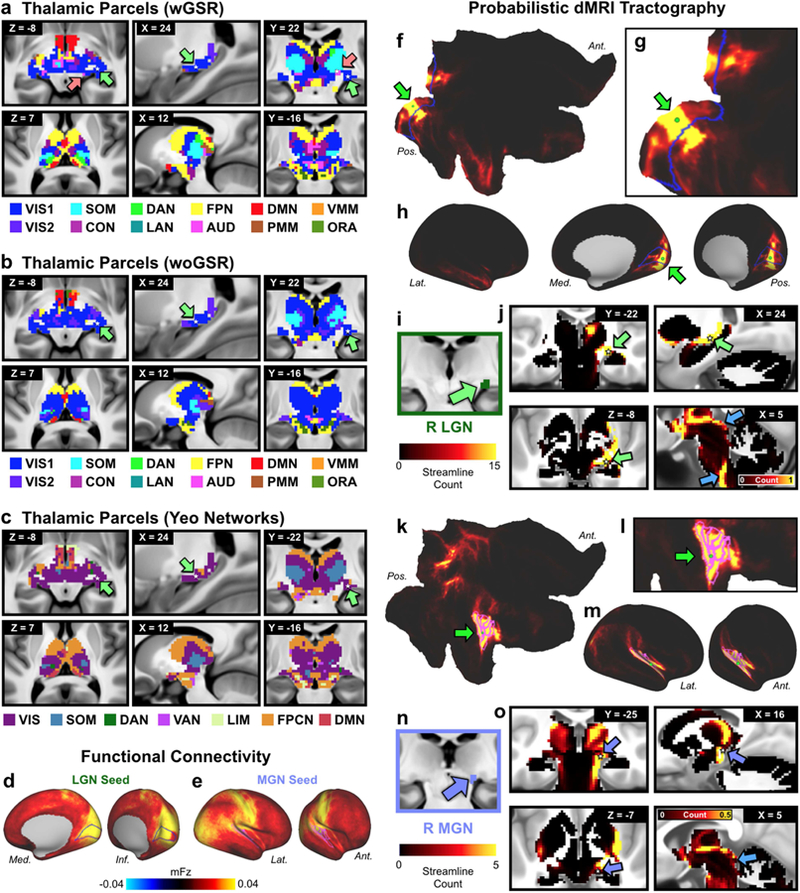
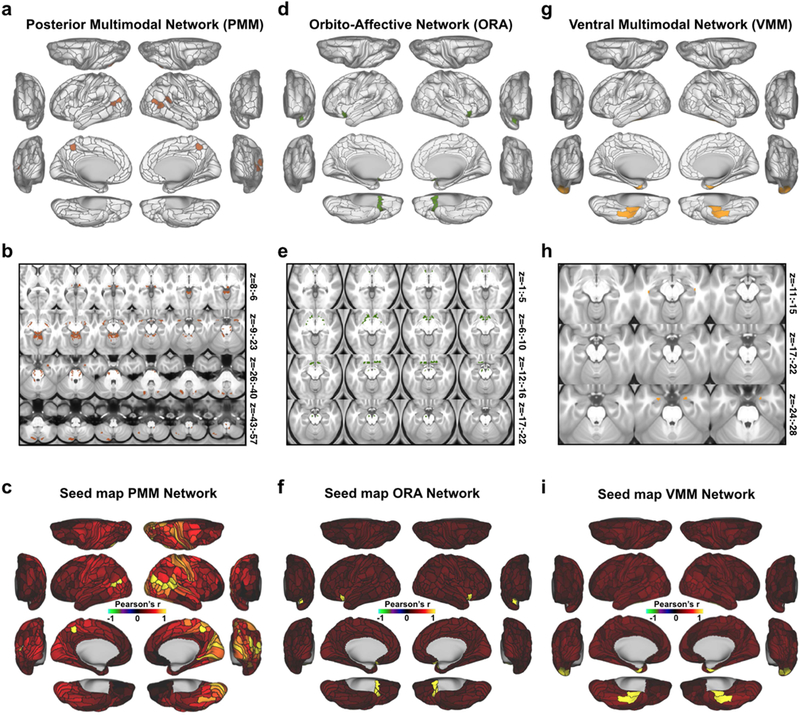
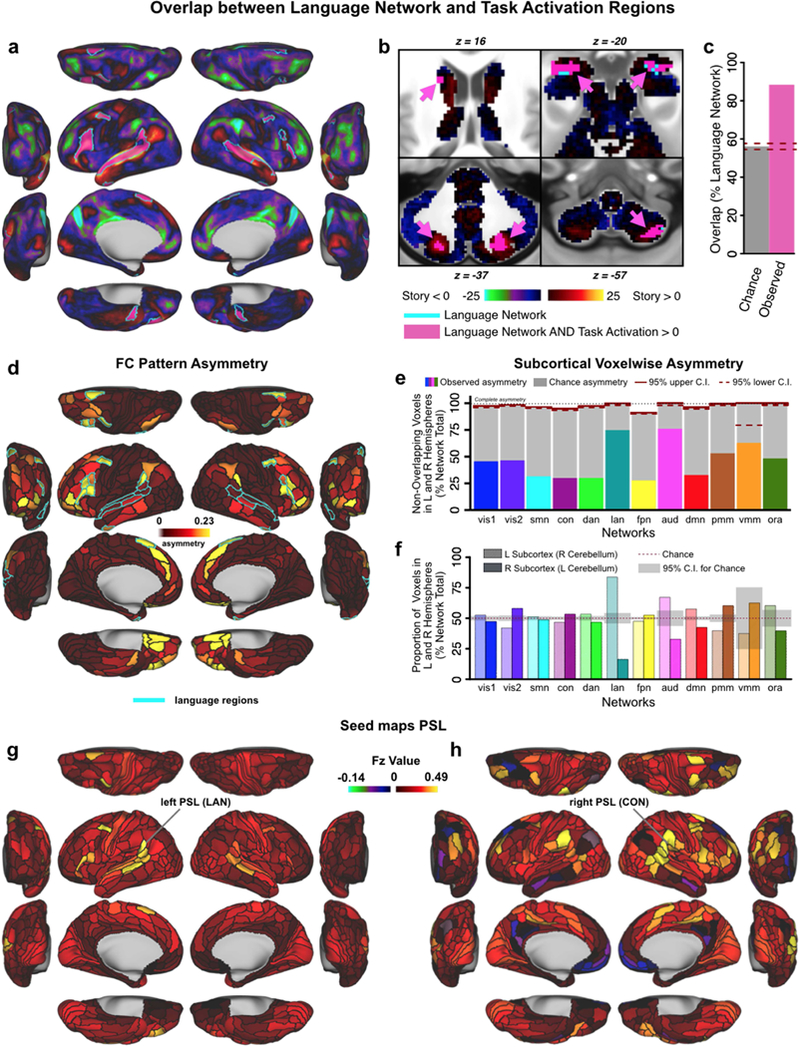
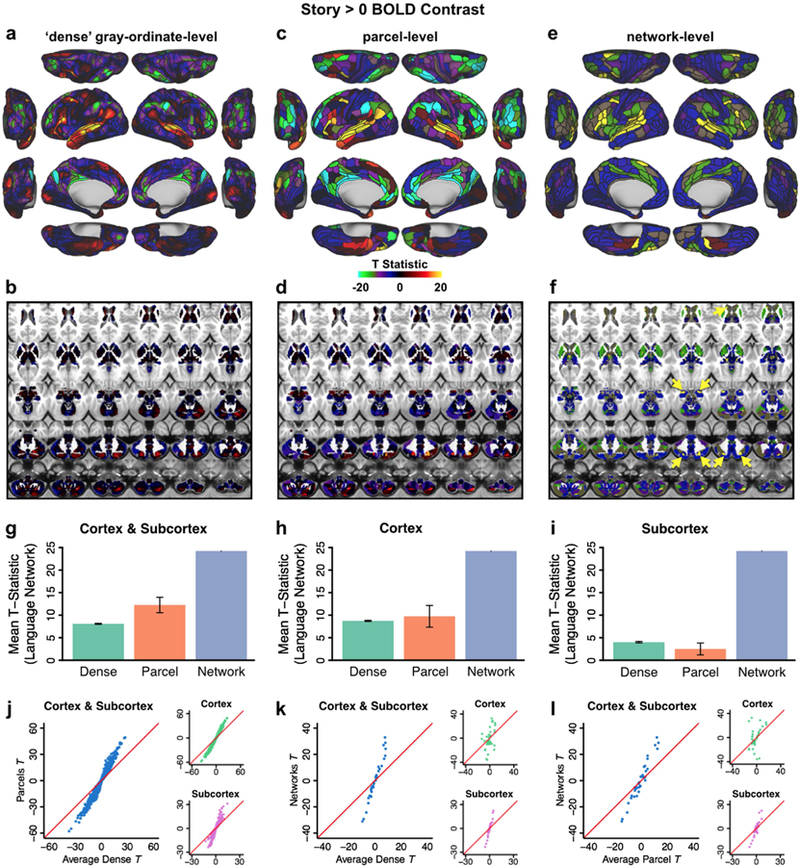
Understanding complex systems such as the human brain requires characterization of the system's architecture across multiple levels of organization - from neurons, to local circuits, to brain regions, and ultimately large-scale brain networks. Here we focus on characterizing the human brain's large-scale network organization, as it provides an overall framework for the organization of all other levels. We developed a highly principled approach to identify cortical network communities at the level of functional systems, calibrating our community detection algorithm using extremely well-established sensory and motor systems as guides. Building on previous network partitions, we replicated and expanded upon well-known and recently-identified networks, including several higher-order cognitive networks such as a left-lateralized language network. We expanded these cortical networks to subcortex, revealing 358 highly-organized subcortical parcels that take part in forming whole-brain functional networks. Notably, the identified subcortical parcels are similar in number to a recent estimate of the number of cortical parcels (360). This whole-brain network atlas - released as an open resource for the neuroscience community - places all brain structures across both cortex and subcortex into a single large-scale functional framework, with the potential to facilitate a variety of studies investigating large-scale functional networks in health and disease.
Keywords: Brain connectivity; Brain networks; Functional MRI; Resting-state functional connectivity.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
References
-
- Barbas H, 2000. Connections underlying the synthesis of cognition, memory, and emotion in primate prefrontal cortices. Brain Res. Bull 52, 319–330. - PubMed
-
- Barch DM, Burgess GC, Harms MP, Petersen SE, Schlaggar BL, Corbetta M, Glasser MF, Curtiss S, Dixit S, Feldt C, Nolan D, Bryant E, Hartley T, Footer O, Bjork JM, Poldrack R, Smith S, Johansen-Berg H, Snyder AZ, Van Essen DC, Consortium, for the WU-Minn HCP, 2013. Function in the human connectome: Task-fMRI and individual differences in behavior. Neuroimage 80, 169–189. - PMC - PubMed
-
- Behrens TEJ, Johansen-Berg H, Woolrich MW, Smith SM, Wheeler-Kingshott CAM, Boulby PA, Barker GJ, Sillery EL, Sheehan K, Ciccarelli O, Thompson AJ, Brady JM, Matthews PM, 2003. Non-invasive mapping of connections between human thalamus and cortex using diffusion imaging. Nat. Neurosci 6, 750–757. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
