

Mechanisms underlying modulation of podocyte TRPC6 channels by suPAR: Role of NADPH oxidases and Src family tyrosine kinases
- PMID: 30293571
- PMCID: PMC6219472
- DOI: 10.1016/j.bbadis.2018.08.007
Mechanisms underlying modulation of podocyte TRPC6 channels by suPAR: Role of NADPH oxidases and Src family tyrosine kinases
Abstract
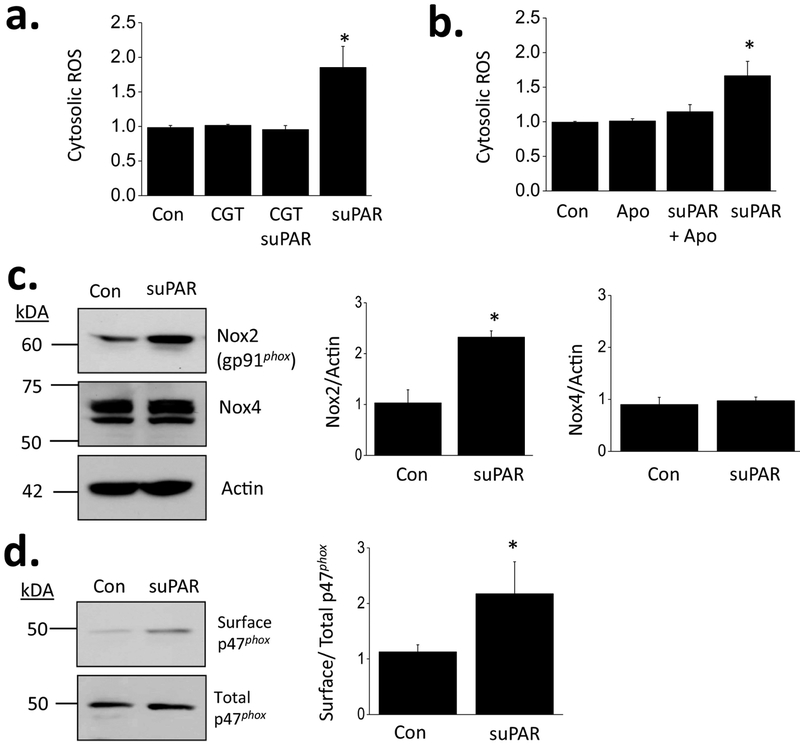
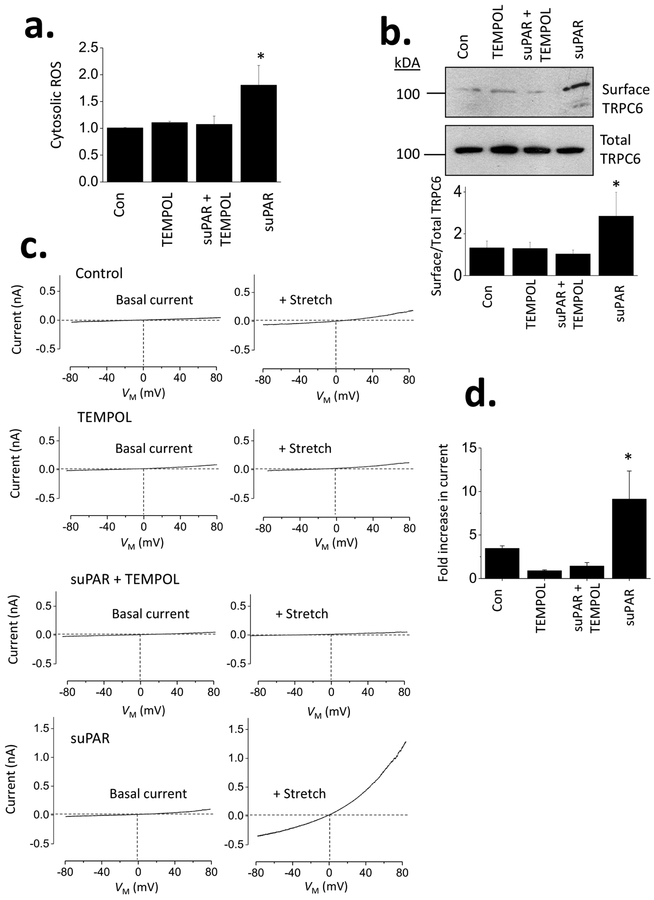
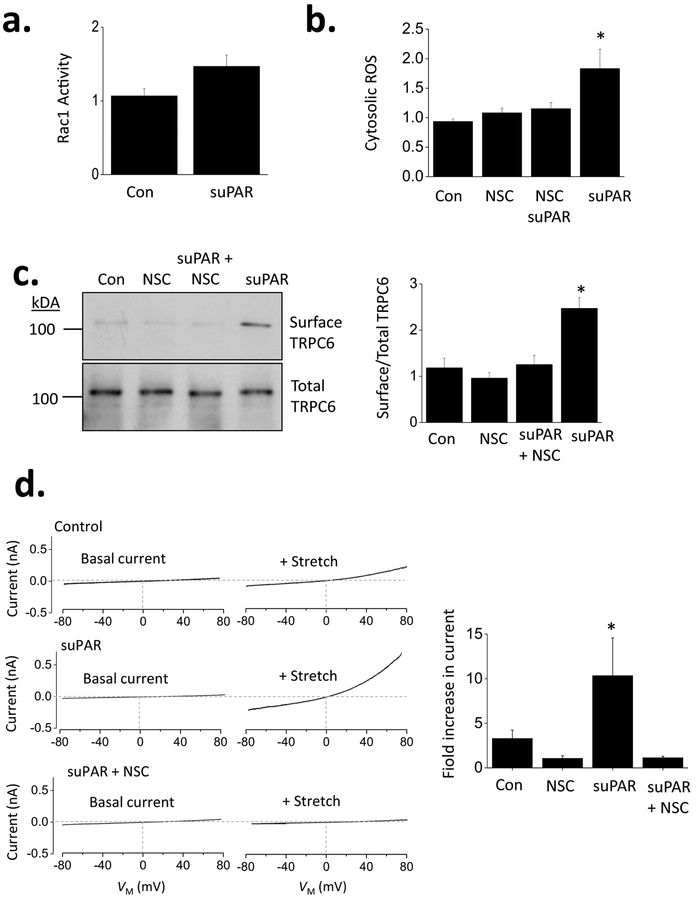
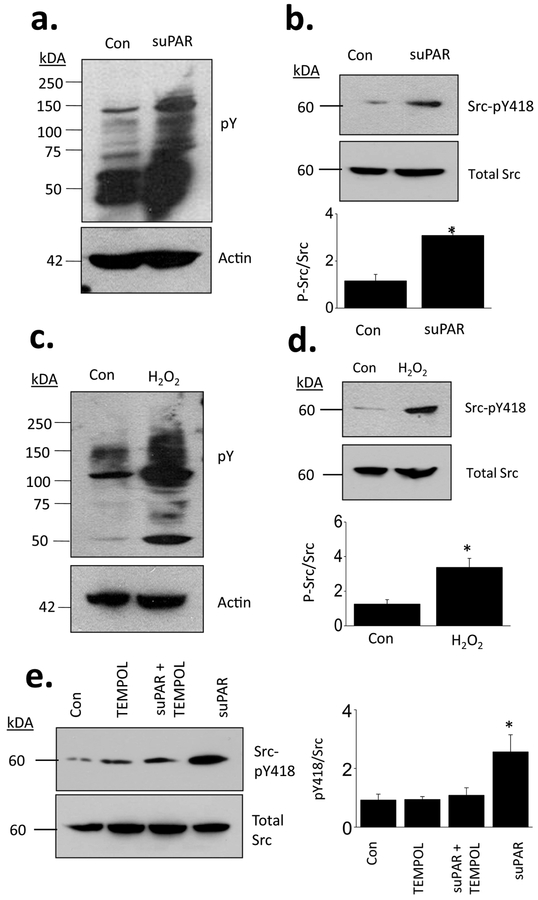
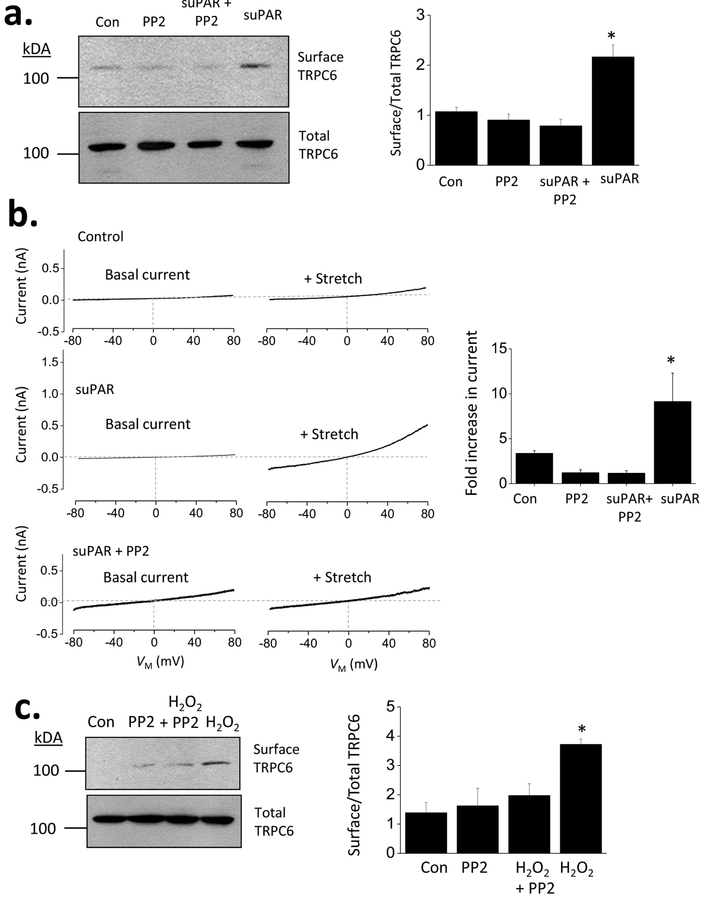
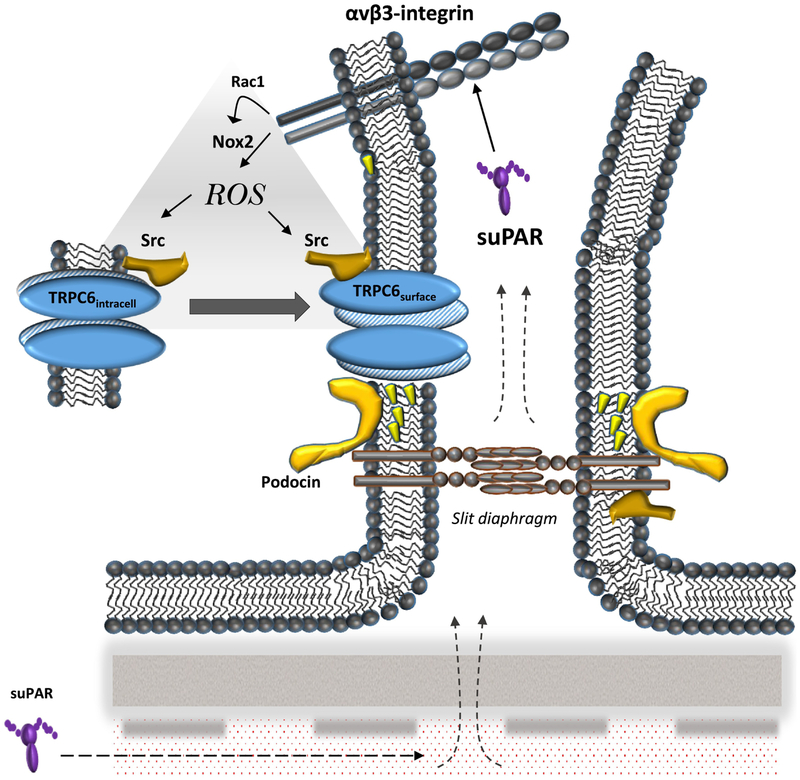
The soluble urokinase receptor (suPAR) has been implicated in the pathogenesis of chronic kidney diseases (CKD) and may function as a circulating "permeability factor" driving primary focal and segmental glomerulosclerosis (FSGS). Here we examined the mechanisms whereby suPAR causes mobilization and increased activation of Ca2+-permeable TRPC6 channels, which are also implicated in FSGS. Treatment of immortalized mouse podocytes with recombinant suPAR for 24 h caused a marked increase in cytosolic reactive oxygen species (ROS) that required signaling through integrins. This effect was associated with increased assembly of active cell surface NADPH oxidase 2 (Nox2) complexes and was blocked by the Nox2 inhibitor apoycynin. Treatment with suPAR also evoked a functionally measurable increase in TRPC6 channels that was blocked by concurrent treatment with the ROS-quencher TEMPOL as well as by inhibition of Rac1, an essential component of active Nox2 complexes. Elevated ROS evoked by exposing cells to suPAR or H2O2 caused a marked increase in the abundance of tyrosine-phosphorylated proteins including Src, and suPAR-evoked Src activation was blocked by TEMPOL. Moreover, mobilization and increased activation of TRPC6 by suPAR or H2O2 was blocked by concurrent exposure to PP2, an inhibitor of Src family tyrosine kinases. These data suggest that suPAR induces oxidative stress in podocytes that in turn drives signaling through Src family kinases to upregulate TRPC6 channels. The combination of oxidative stress and altered Ca2+ signaling may contribute to loss of podocytes and progression of various forms of CKD.
Keywords: Chronic kidney disease; Integrins; Reactive oxygen species; Src; TRPC6; suPAR.
Copyright © 2018 Elsevier B.V. All rights reserved.
Figures
References
-
- Montuori N, Carriero MV, Salzano S, Rossi G, Ragno P. The cleavage of the urokinase receptor regulates its multiple functions. J BiolChem. 2002. 277(49):46932–9. - PubMed
-
- Desmedt S, Desmedt V, Delanghe JR, Speeckaert R, Speeckaert MM. The intriguing role of soluble urokinase receptor in inflammatory diseases. Crit Rev Clin Lab Sci. 2017. 54(2):117–133. - PubMed
-
- Zeier M, Reiser J. suPAR and chronic kidney disease-a podocyte story. Pflugers Arch. 2017. 469(7–8):1017–1020. - PubMed
-
- Blosser CD, Bloom RD. Recurrent glomerular disease after kidney transplantation. Curr Opin Nephrol Hypertens. 2017. 26(6):501–508. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
