

Genome-wide Identification of Structure-Forming Repeats as Principal Sites of Fork Collapse upon ATR Inhibition
- PMID: 30293786
- PMCID: PMC6407864
- DOI: 10.1016/j.molcel.2018.08.047
Genome-wide Identification of Structure-Forming Repeats as Principal Sites of Fork Collapse upon ATR Inhibition
Abstract
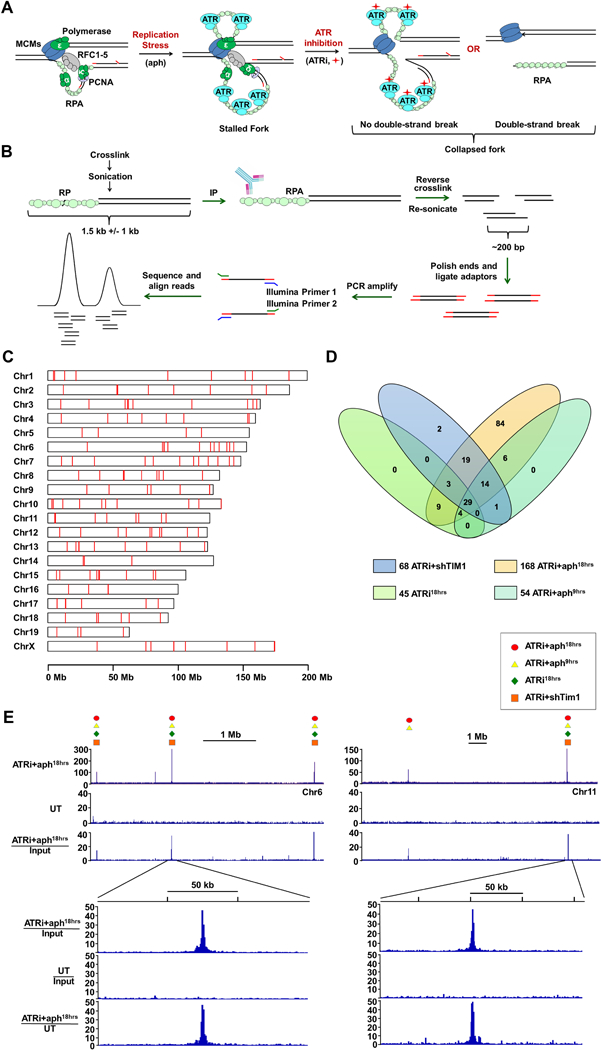
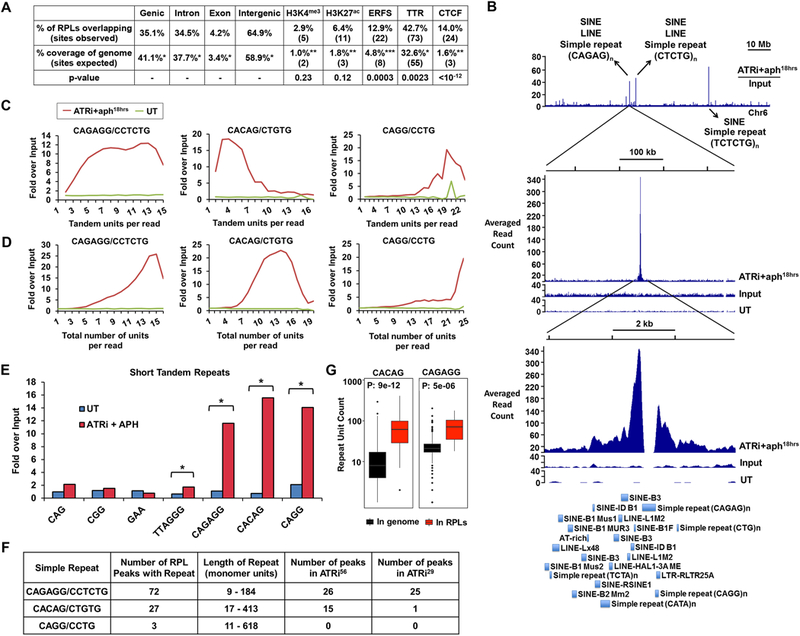
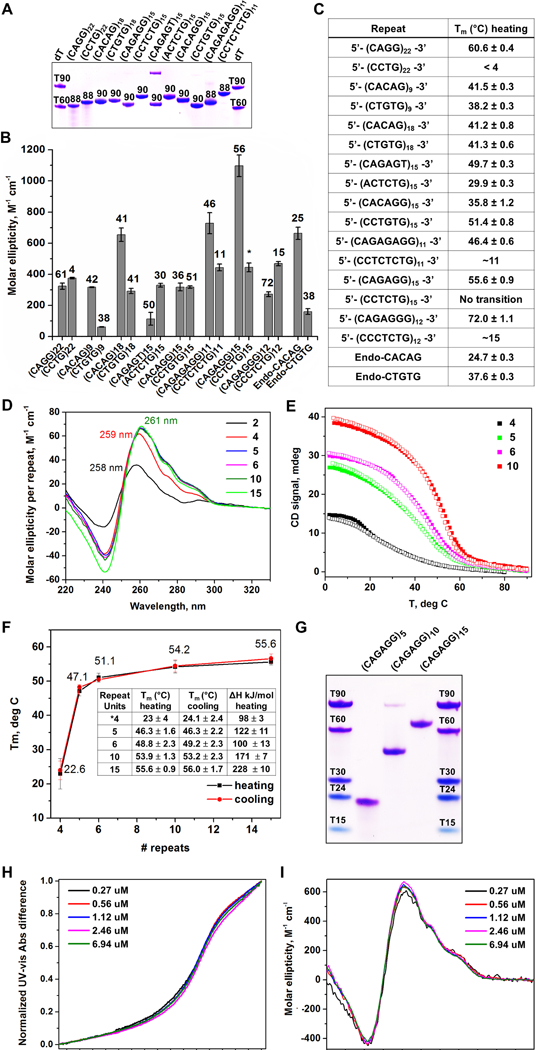
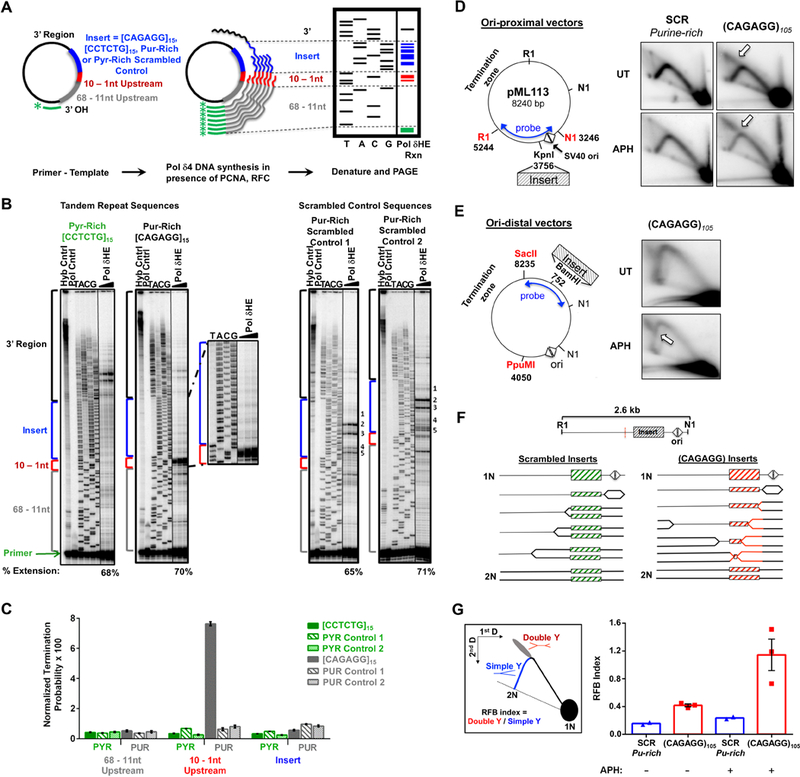
DNA polymerase stalling activates the ATR checkpoint kinase, which in turn suppresses fork collapse and breakage. Herein, we describe use of ATR inhibition (ATRi) as a means to identify genomic sites of problematic DNA replication in murine and human cells. Over 500 high-resolution ATR-dependent sites were ascertained using two distinct methods: replication protein A (RPA)-chromatin immunoprecipitation (ChIP) and breaks identified by TdT labeling (BrITL). The genomic feature most strongly associated with ATR dependence was repetitive DNA that exhibited high structure-forming potential. Repeats most reliant on ATR for stability included structure-forming microsatellites, inverted retroelement repeats, and quasi-palindromic AT-rich repeats. Notably, these distinct categories of repeats differed in the structures they formed and their ability to stimulate RPA accumulation and breakage, implying that the causes and character of replication fork collapse under ATR inhibition can vary in a DNA-structure-specific manner. Collectively, these studies identify key sources of endogenous replication stress that rely on ATR for stability.
Keywords: AT-rich; ATR; DNA damage; DNA double-strand breaks; RPA; hairpin; inverted repeats; microsatellite; replication fork collapse; short tandem repeats.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
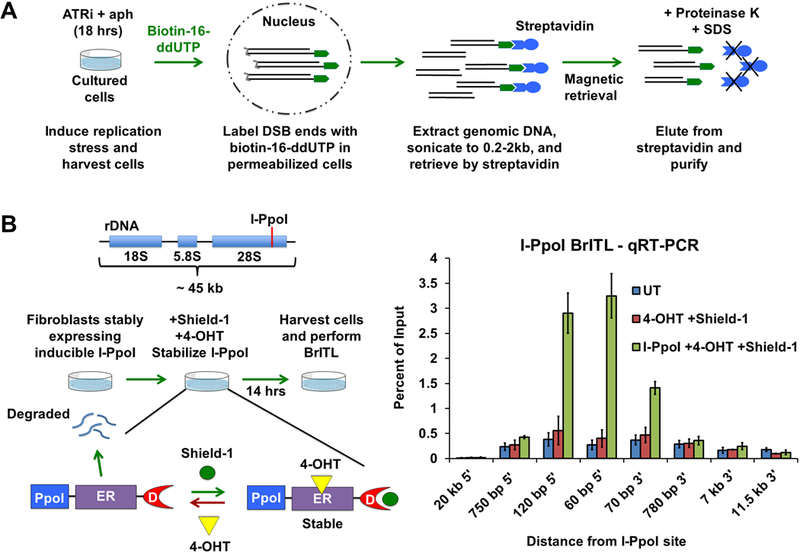
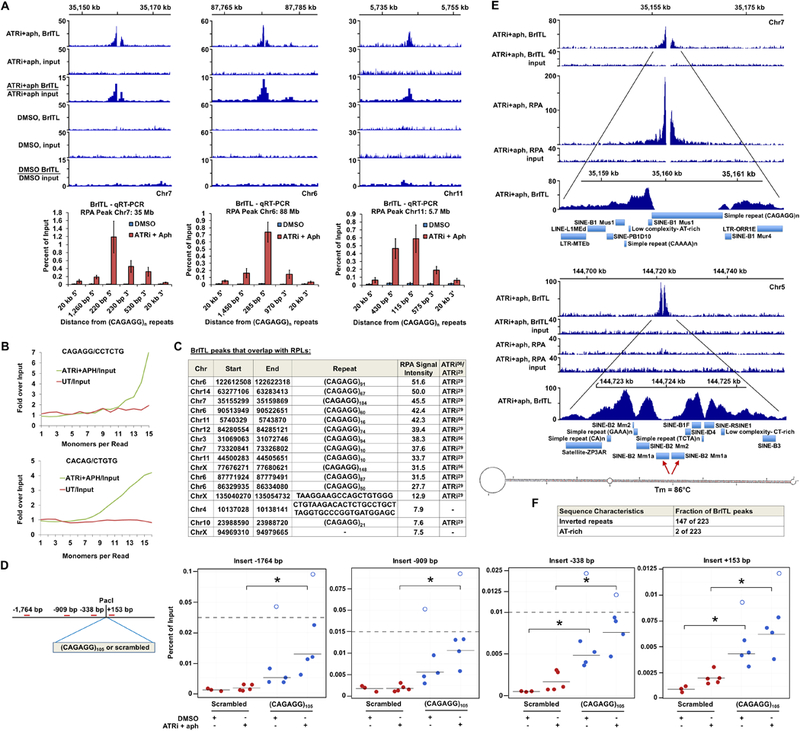
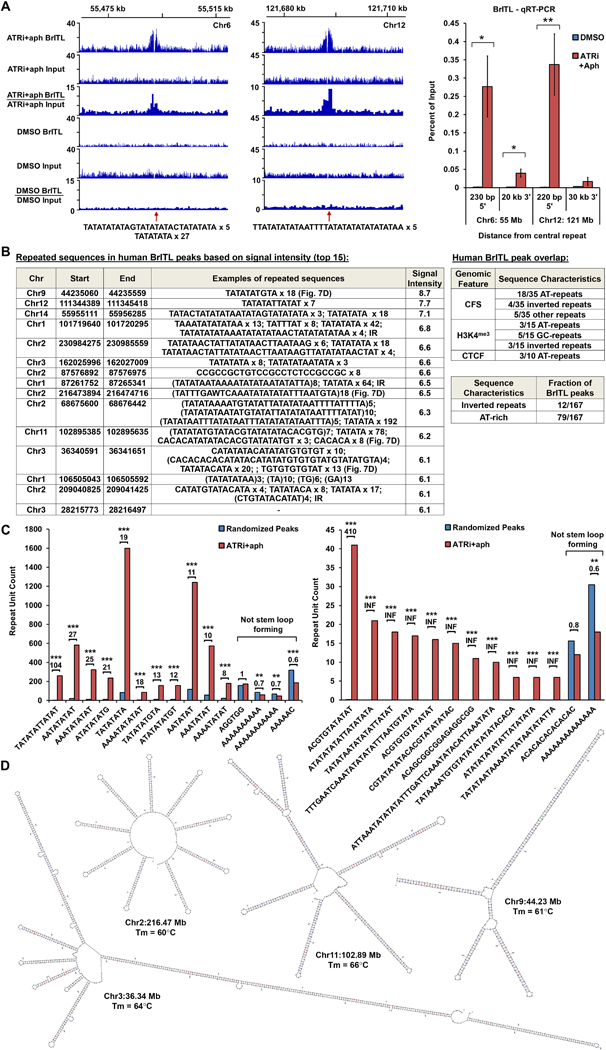
Comment in
-
Getting a Genomic View of DNA Replication Stress.Mol Cell. 2018 Oct 18;72(2):201-203. doi: 10.1016/j.molcel.2018.10.001. Mol Cell. 2018. PMID: 30340018
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
