

Neural Coding of Leg Proprioception in Drosophila
- PMID: 30293823
- PMCID: PMC6481666
- DOI: 10.1016/j.neuron.2018.09.009
Neural Coding of Leg Proprioception in Drosophila
Abstract
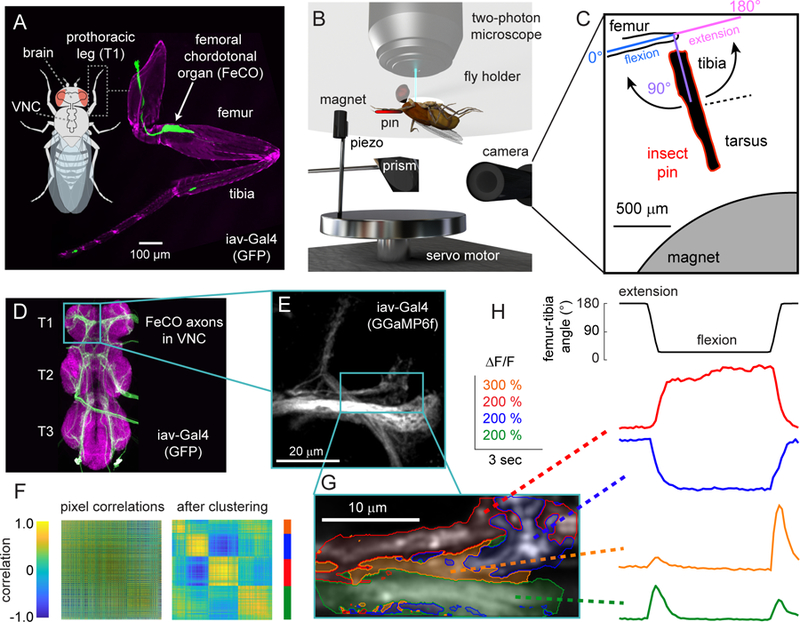
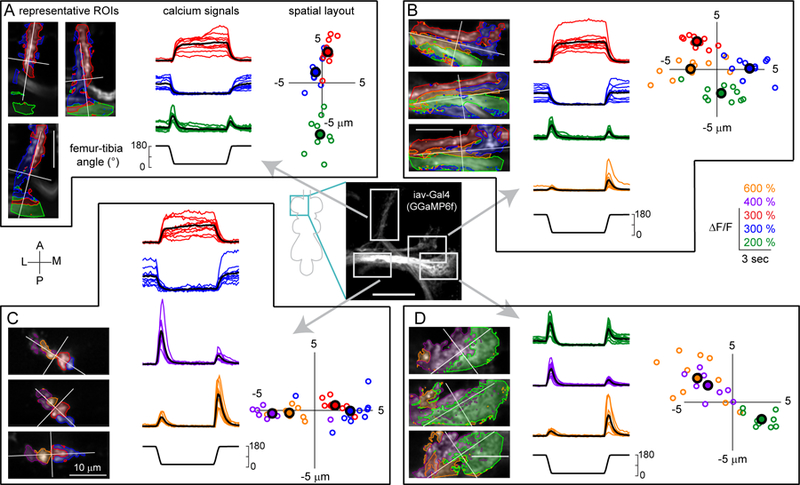
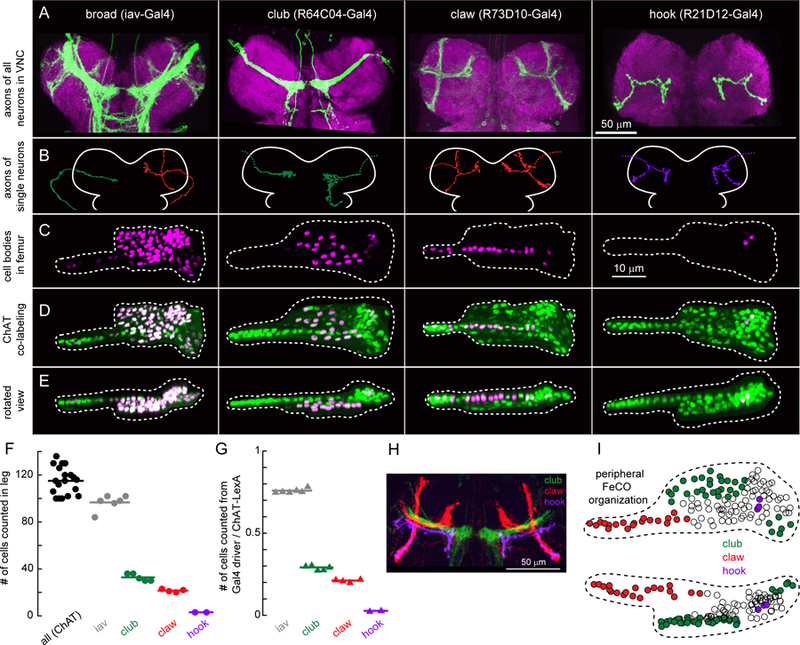
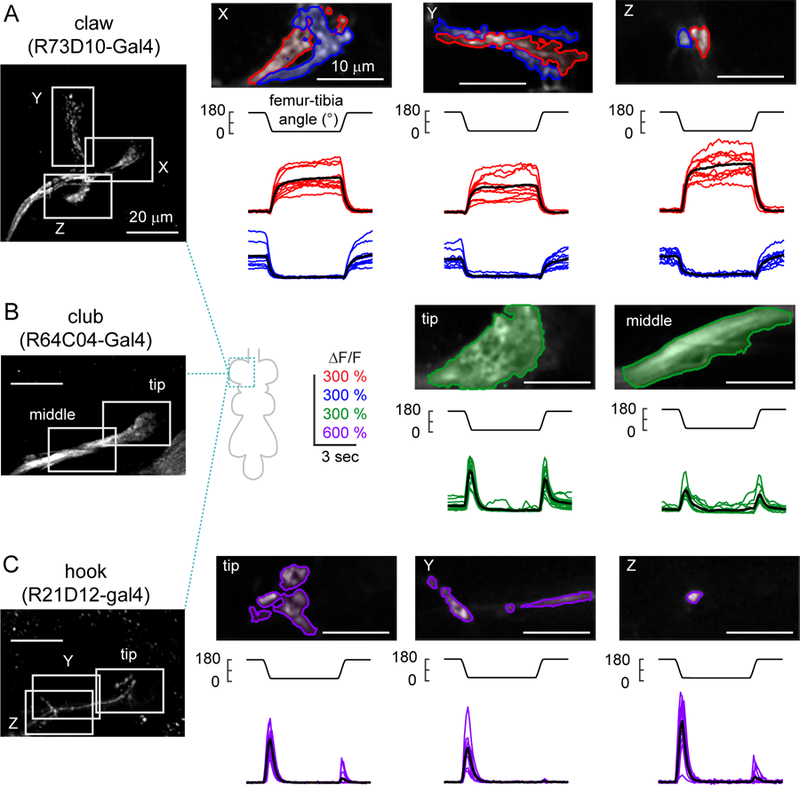
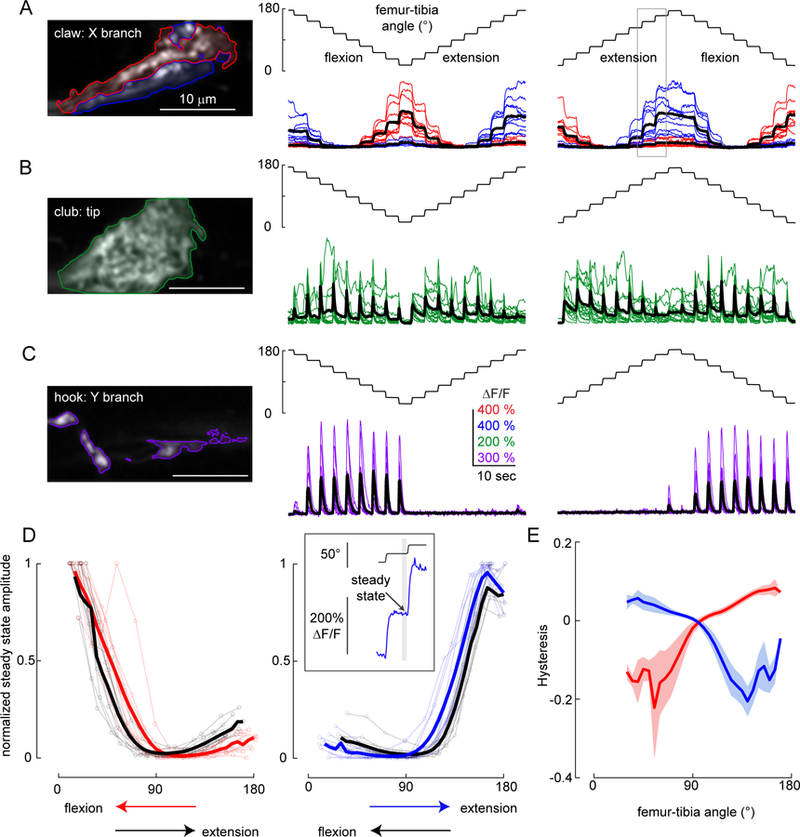
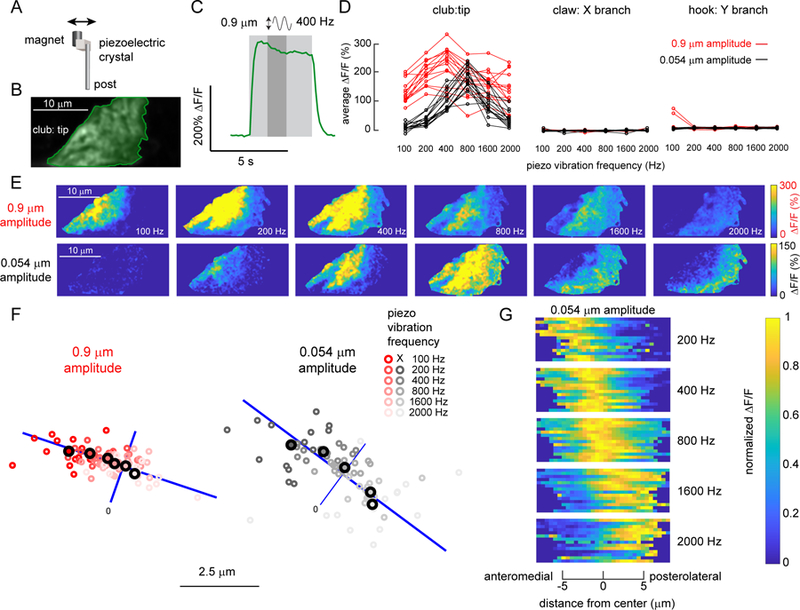
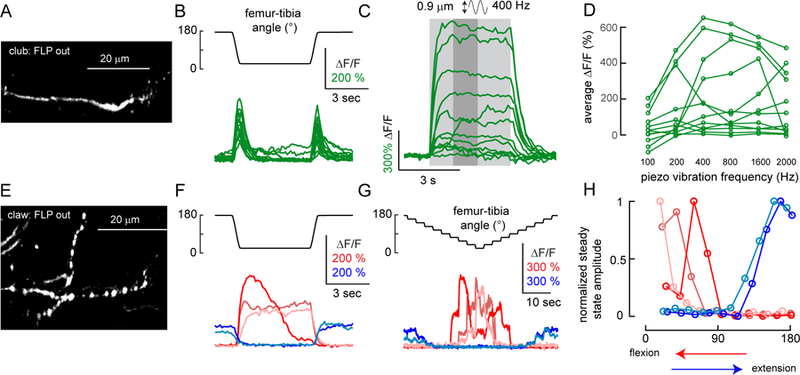
Animals rely on an internal sense of body position and movement to effectively control motor behavior. This sense of proprioception is mediated by diverse populations of mechanosensory neurons distributed throughout the body. Here, we investigate neural coding of leg proprioception in Drosophila, using in vivo two-photon calcium imaging of proprioceptive sensory neurons during controlled movements of the fly tibia. We found that the axons of leg proprioceptors are organized into distinct functional projections that contain topographic representations of specific kinematic features. Using subclass-specific genetic driver lines, we show that one group of axons encodes tibia position (flexion/extension), another encodes movement direction, and a third encodes bidirectional movement and vibration frequency. Overall, our findings reveal how proprioceptive stimuli from a single leg joint are encoded by a diverse population of sensory neurons, and provide a framework for understanding how proprioceptive feedback signals are used by motor circuits to coordinate the body.
Keywords: Drosophila; chordotonal; mechanosensation; mechanosensory; motor control; neural coding; proprioception; proprioceptive; proprioceptor; two-photon imaging.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests
The authors declare no competing interests.
Figures
Similar articles
-
Proprioception, the regulator of motor function.BMB Rep. 2021 Aug;54(8):393-402. doi: 10.5483/BMBRep.2021.54.8.052. BMB Rep. 2021. PMID: 34078529 Free PMC article. Review.
-
Central processing of leg proprioception in Drosophila.Elife. 2020 Dec 2;9:e60299. doi: 10.7554/eLife.60299. Elife. 2020. PMID: 33263281 Free PMC article.
-
Functional architecture of neural circuits for leg proprioception in Drosophila.Curr Biol. 2021 Dec 6;31(23):5163-5175.e7. doi: 10.1016/j.cub.2021.09.035. Epub 2021 Oct 11. Curr Biol. 2021. PMID: 34637749 Free PMC article.
-
Biomechanical origins of proprioceptor feature selectivity and topographic maps in the Drosophila leg.Neuron. 2023 Oct 18;111(20):3230-3243.e14. doi: 10.1016/j.neuron.2023.07.009. Epub 2023 Aug 9. Neuron. 2023. PMID: 37562405 Free PMC article.
-
The making of a proprioceptor: a tale of two identities.Trends Neurosci. 2023 Dec;46(12):1083-1094. doi: 10.1016/j.tins.2023.09.008. Epub 2023 Oct 17. Trends Neurosci. 2023. PMID: 37858440 Review.
Cited by
-
Proprioception, the regulator of motor function.BMB Rep. 2021 Aug;54(8):393-402. doi: 10.5483/BMBRep.2021.54.8.052. BMB Rep. 2021. PMID: 34078529 Free PMC article. Review.
-
A size principle for recruitment of Drosophila leg motor neurons.Elife. 2020 Jun 3;9:e56754. doi: 10.7554/eLife.56754. Elife. 2020. PMID: 32490810 Free PMC article.
-
Emergence of social cluster by collective pairwise encounters in Drosophila.Elife. 2020 Jan 21;9:e51921. doi: 10.7554/eLife.51921. Elife. 2020. PMID: 31959283 Free PMC article.
-
Global change in brain state during spontaneous and forced walk in Drosophila is composed of combined activity patterns of different neuron classes.Elife. 2023 Apr 17;12:e85202. doi: 10.7554/eLife.85202. Elife. 2023. PMID: 37067152 Free PMC article.
-
Central processing of leg proprioception in Drosophila.Elife. 2020 Dec 2;9:e60299. doi: 10.7554/eLife.60299. Elife. 2020. PMID: 33263281 Free PMC article.
References
-
- Ahrens MB, Orger MB, Robson DN, Li JM, and Keller PJ (2013). Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nat Methods 10, 413–420. - PubMed
-
- Bässler U (1988). Functional principles of pattern generation for walking movements of stick insect forelegs: the role of the femoral chordotonal organ afferences. Journal of Experimental Biology 136, 125–147.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
