

OmpR-Mediated Transcriptional Regulation and Function of Two Heme Receptor Proteins of Yersinia enterocolitica Bio-Serotype 2/O:9
- PMID: 30294593
- PMCID: PMC6158557
- DOI: 10.3389/fcimb.2018.00333
OmpR-Mediated Transcriptional Regulation and Function of Two Heme Receptor Proteins of Yersinia enterocolitica Bio-Serotype 2/O:9
Abstract
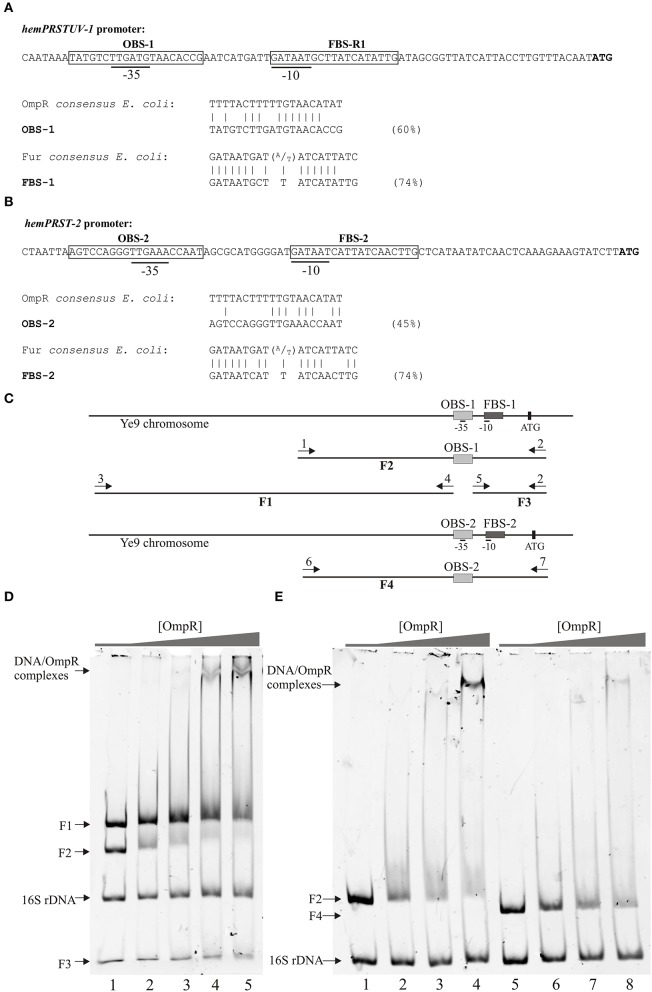
We show that Yersinia enterocolitica strain Ye9 (bio-serotype 2/O:9) utilizes heme-containing molecules as an iron source. The Ye9 genome contains two multigenic clusters, hemPRSTUV-1 and hemPRST-2, encoding putative heme receptors HemR1 and HemR2, that share 62% amino acid identity. Expression of these proteins in an Escherichia coli mutant defective in heme biosynthesis allowed this strain to use hemin and hemoglobin as a source of porphyrin. The hemPRSTUV-1 and hemPRST-2 clusters are organized as operons, expressed from the phem-1 and weaker phem-2 promoters, respectively. Expression of both operons is negatively regulated by iron and the iron-responsive transcriptional repressor Fur. In addition, OmpR, the response regulator of two component system (TCSs) EnvZ/OmpR, represses transcription of both operons through interaction with binding sequences overlapping the -35 region of their promoters. Western blot analysis of the level of HemR1 in ompR, fur, and ompRfur mutants, showed an additive effect of these mutations, indicating that OmpR may regulate HemR expression independently of Fur. However, the effect of OmpR on the activity of the phem-1 promoter and on HemR1 production was observed in both iron-depleted and iron-replete conditions, i.e., when Fur represses the iron-regulated promoter. In addition, a hairpin RNA thermometer, composed of four uracil residues (FourU) that pair with the ribosome-binding site in the 5'-untranslated region (5'-UTR) of hemR1 was predicted by in silico analysis. However, thermoregulated expression of HemR1 could not be demonstrated. Taken together, these data suggest that Fur and OmpR control iron/heme acquisition via a complex mechanism based on negative regulation of hemR1 and hemR2 at the transcriptional level. This interplay could fine-tune the level of heme receptor proteins to allow Y. enterocolitica to fulfill its iron/heme requirements without over-accumulation, which might be important for pathogenic growth within human hosts.
Keywords: Fur; HemR1; HemR2; OmpR; Yersinia enterocolitica.
Figures







Similar articles
-
The Regulator OmpR in Yersinia enterocolitica Participates in Iron Homeostasis by Modulating Fur Level and Affecting the Expression of Genes Involved in Iron Uptake.Int J Mol Sci. 2021 Feb 2;22(3):1475. doi: 10.3390/ijms22031475. Int J Mol Sci. 2021. PMID: 33540627 Free PMC article.
-
The Role of OmpR in the Expression of Genes of the KdgR Regulon Involved in the Uptake and Depolymerization of Oligogalacturonides in Yersinia enterocolitica.Front Cell Infect Microbiol. 2017 Aug 15;7:366. doi: 10.3389/fcimb.2017.00366. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28861396 Free PMC article.
-
OmpR negatively regulates expression of invasin in Yersinia enterocolitica.Microbiology (Reading). 2007 Aug;153(Pt 8):2416-2425. doi: 10.1099/mic.0.2006/003202-0. Microbiology (Reading). 2007. PMID: 17660406
-
Role and regulation of heme iron acquisition in gram-negative pathogens.Front Cell Infect Microbiol. 2013 Oct 8;3:55. doi: 10.3389/fcimb.2013.00055. eCollection 2013. Front Cell Infect Microbiol. 2013. PMID: 24116354 Free PMC article. Review.
-
Iron-regulated surface determinants (Isd) of Staphylococcus aureus: stealing iron from heme.Microbes Infect. 2004 Apr;6(4):390-7. doi: 10.1016/j.micinf.2003.12.008. Microbes Infect. 2004. PMID: 15101396 Review.
Cited by
-
Two-component systems regulate bacterial virulence in response to the host gastrointestinal environment and metabolic cues.Virulence. 2022 Dec;13(1):1666-1680. doi: 10.1080/21505594.2022.2127196. Virulence. 2022. PMID: 36128741 Free PMC article. Review.
-
Urease Expression in Pathogenic Yersinia enterocolitica Strains of Bio-Serotypes 2/O:9 and 1B/O:8 Is Differentially Regulated by the OmpR Regulator.Front Microbiol. 2020 Apr 8;11:607. doi: 10.3389/fmicb.2020.00607. eCollection 2020. Front Microbiol. 2020. PMID: 32322248 Free PMC article.
-
Biotype Determines Survival of Yersinia enterocolitica in Red Blood Cell Concentrates.Int J Mol Sci. 2025 Jun 16;26(12):5775. doi: 10.3390/ijms26125775. Int J Mol Sci. 2025. PMID: 40565236 Free PMC article.
-
The Regulator OmpR in Yersinia enterocolitica Participates in Iron Homeostasis by Modulating Fur Level and Affecting the Expression of Genes Involved in Iron Uptake.Int J Mol Sci. 2021 Feb 2;22(3):1475. doi: 10.3390/ijms22031475. Int J Mol Sci. 2021. PMID: 33540627 Free PMC article.
-
The Regulatory Circuit Underlying Downregulation of a Type III Secretion System in Yersinia enterocolitica by Transcription Factor OmpR.Int J Mol Sci. 2022 Apr 26;23(9):4758. doi: 10.3390/ijms23094758. Int J Mol Sci. 2022. PMID: 35563149 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources