

It's complicated… m6A-dependent regulation of gene expression in cancer
- PMID: 30296493
- PMCID: PMC6401303
- DOI: 10.1016/j.bbagrm.2018.09.010
It's complicated… m6A-dependent regulation of gene expression in cancer
Abstract
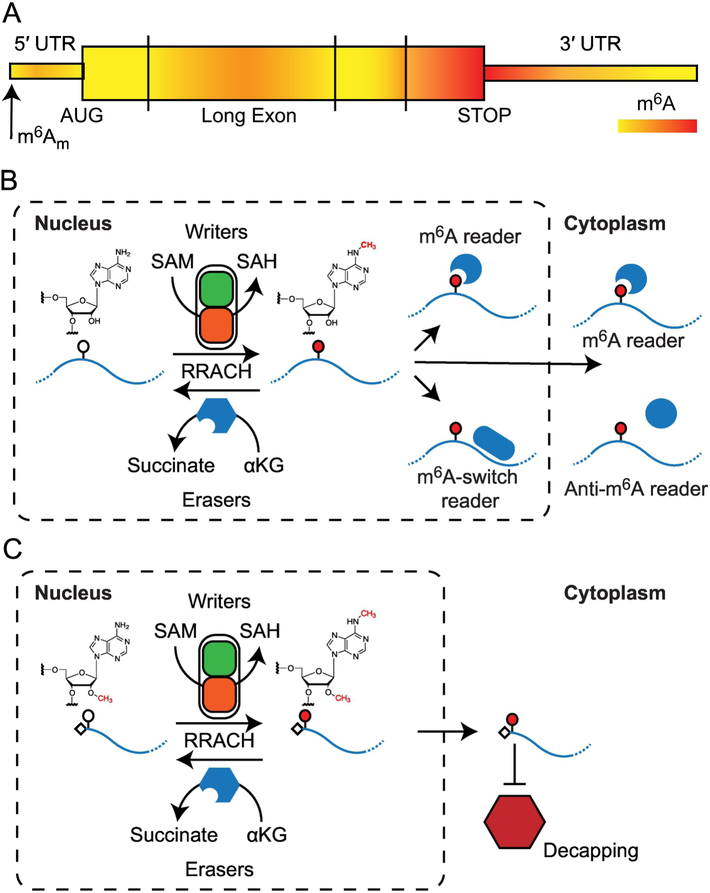
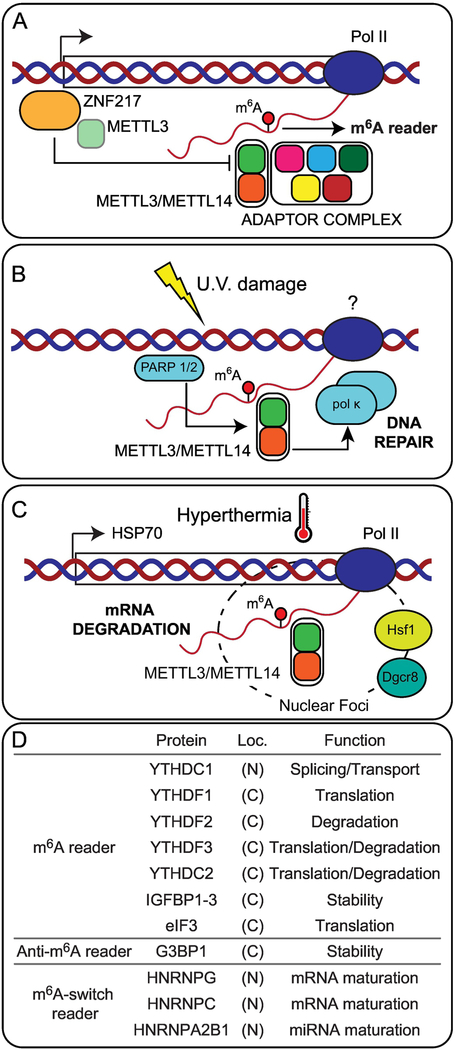
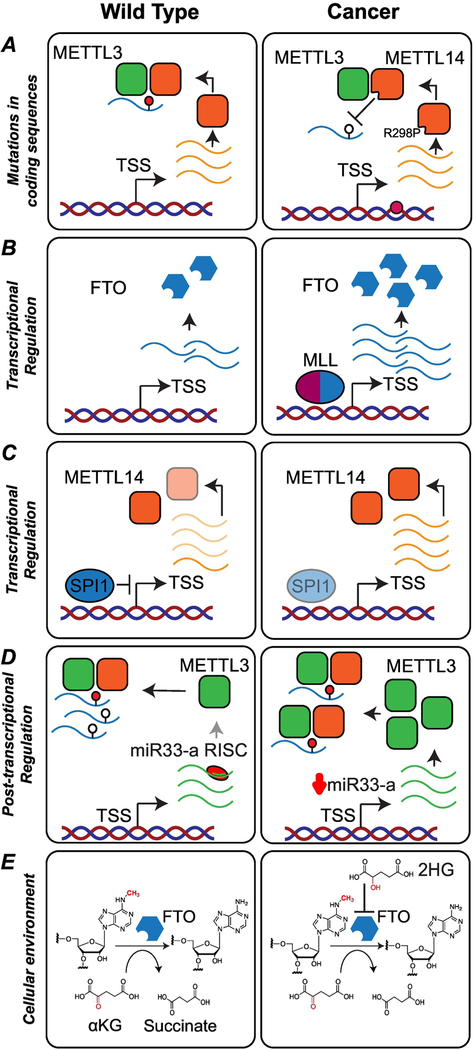
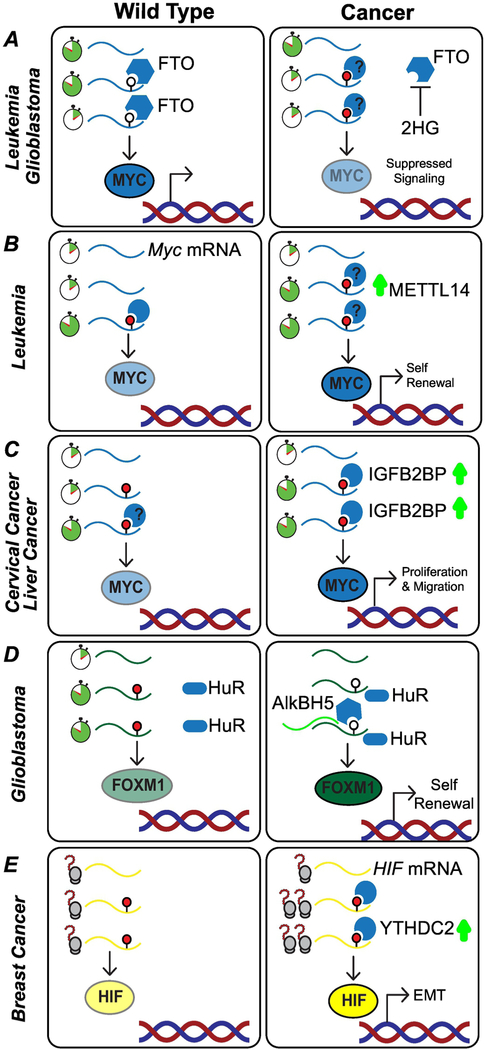
Cellular function relies on multiple pathways that are coordinated to ensure the proper execution of gene expression networks. Failure to coordinate the multiple programs active in the cell can have catastrophic consequences and lead to diseases such as cancer. At the post-transcriptional level, RNA modifications play important roles in the regulation of gene expression. N6-methyladenosine (m6A) is the most abundant internal messenger RNA (mRNA) modification and has gained increasing interest in the last few years as a dynamic regulator of RNA metabolism. Modifications regulate all stages of the RNA life cycle, from transcription to decay. Recent studies have pointed to the role of RNA methylation in cancer initiation and progression, and aberrant modification has served as a biomarker of early-stage diagnosis in several cancers. Here, we review the regulation of m6A, disruptions to methylation-dependent pathways that influence carcinogenesis, and potential avenues for m6A-related therapeutic strategies.
Keywords: Cancer; Epitranscriptome; Gene regulation; m(6)A.
Published by Elsevier B.V.
Figures
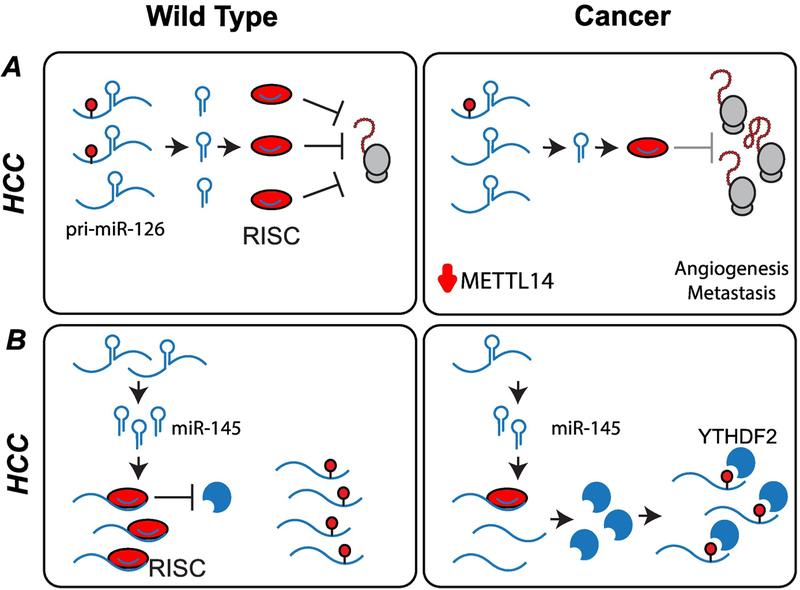
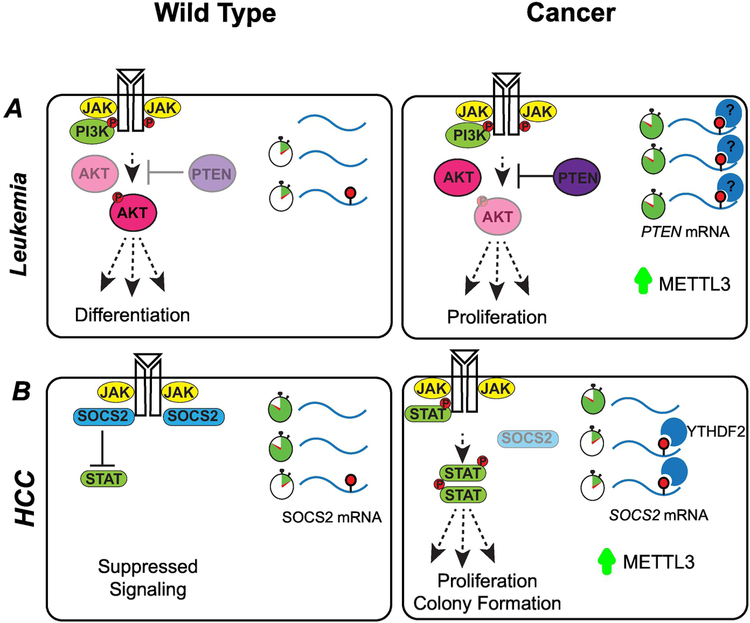
References
-
- Motorin Y, Helm M. RNA nucleotide methylation. Wiley Interdiscip Rev RNA. 2011. October;2(5):611–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
