

Identification of Anti-Melanogenesis Constituents from Morus alba L. Leaves
- PMID: 30297610
- PMCID: PMC6222840
- DOI: 10.3390/molecules23102559
Identification of Anti-Melanogenesis Constituents from Morus alba L. Leaves
Abstract
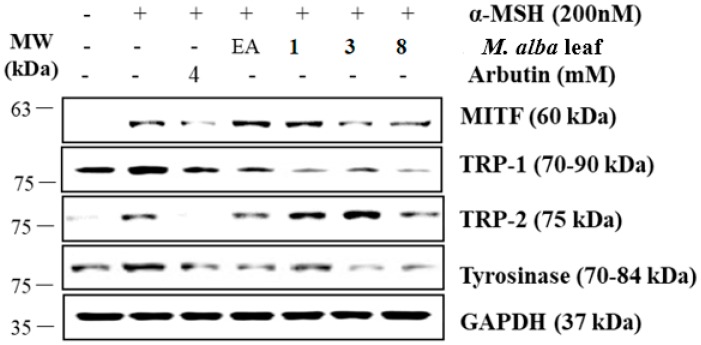
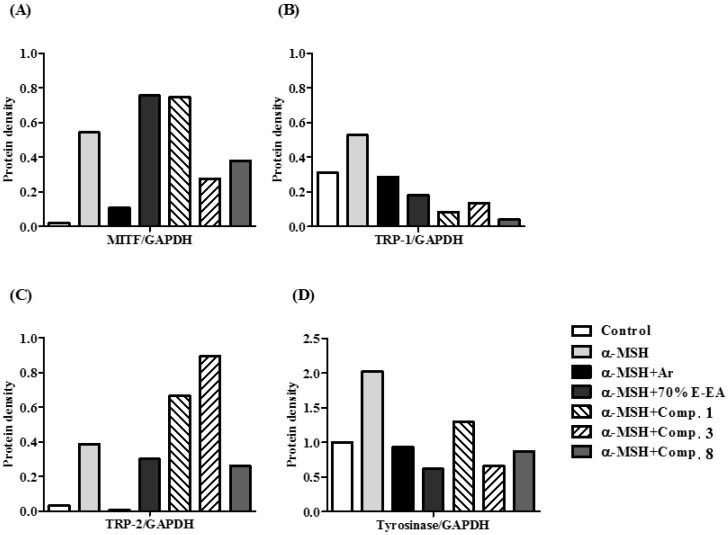
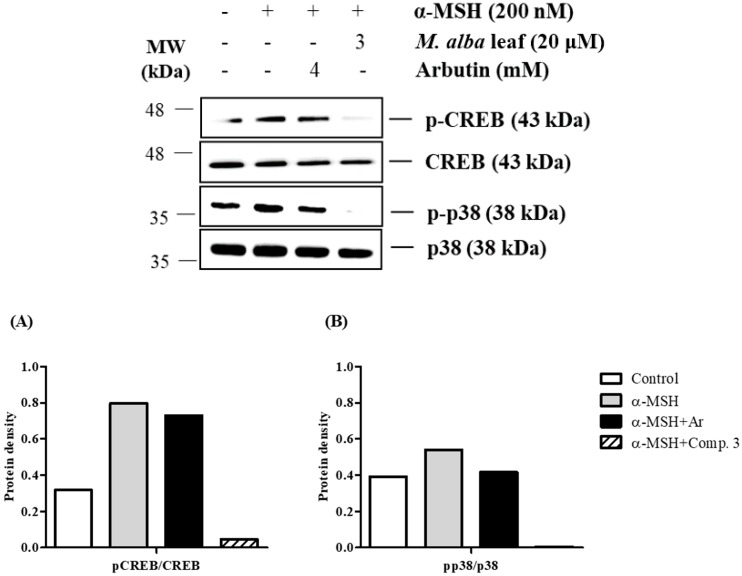
The individual parts of Morus alba L. including root bark, branches, leaves, and fruits are used as a cosmetic ingredient in many Asian countries. This study identified several anti-melanogenesis constituents in a 70% ethanol extract of M. alba leaves. The ethyl acetate fraction of the initial ethanol extract decreased the activity of tyrosinase, a key enzyme in the synthetic pathway of melanin. Twelve compounds were isolated from this fraction and their structures were identified based on spectroscopic spectra. Then, the authors investigated the anti-melanogenesis effects of the isolated compounds in B16-F10 mouse melanoma cells. Compounds 3 and 8 significantly inhibited not only melanin production but also intracellular tyrosinase activity in alpha-melanocyte-stimulating-hormone (α-MSH)-induced B16-F10 cells in a dose-dependent manner. These same compounds also inhibited melanogenesis-related protein expression such as microphthalmia-associated transcription factor (MITF), tyrosinase, and tyrosinase-related protein-1 (TRP-1). Compound 3 modulated the cAMP-responsive element-binding protein (CREB) and p38 signaling pathways in α-MSH-activated B16-F10 melanoma cells, which resulted in the anti-melanogenesis effects. These results suggest that compound 3, isolated from M. alba leaves, could be used to inhibit melanin production via the regulation of melanogenesis-related protein expression.
Keywords: MITF; Morus alba leaf; TRP-1; TRP-2; anti-melanogenesis; tyrosinase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
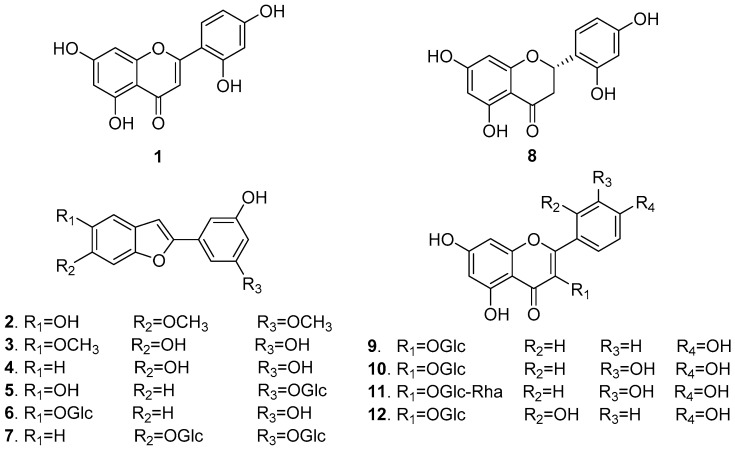
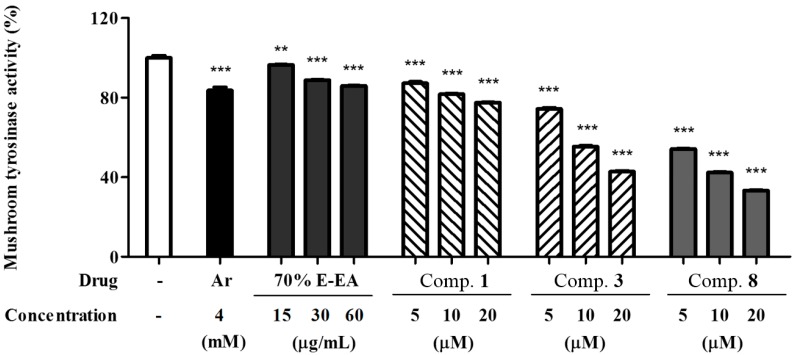
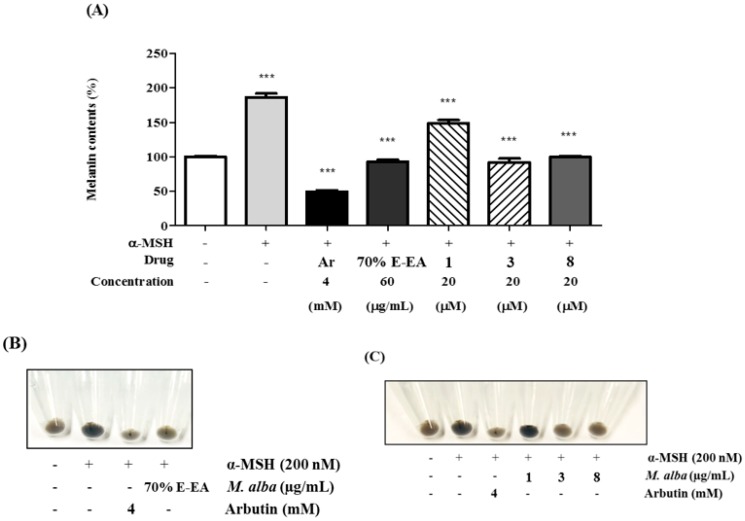
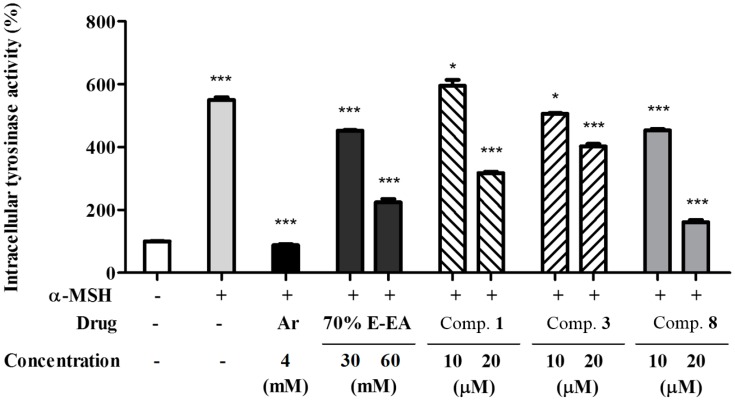
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
