

Matrix mechanical plasticity regulates cancer cell migration through confining microenvironments
- PMID: 30297715
- PMCID: PMC6175826
- DOI: 10.1038/s41467-018-06641-z
Matrix mechanical plasticity regulates cancer cell migration through confining microenvironments
Abstract
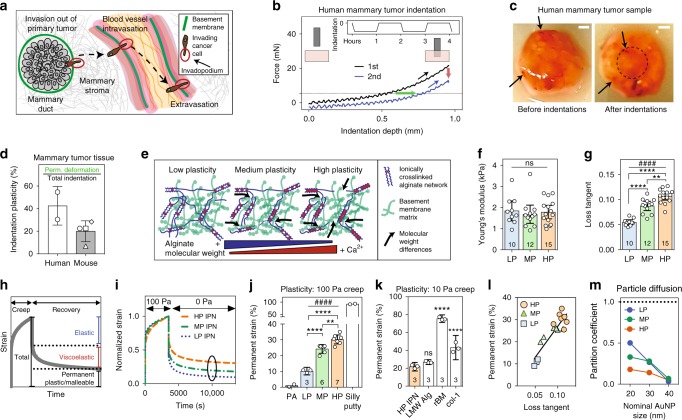
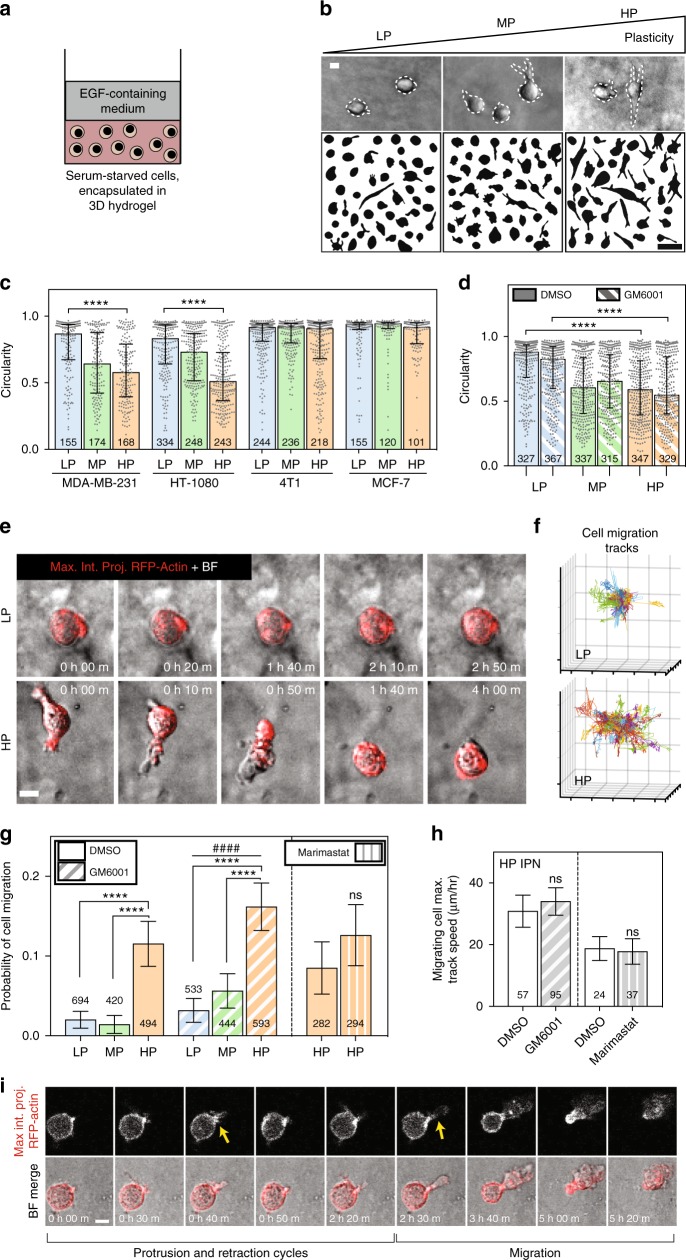
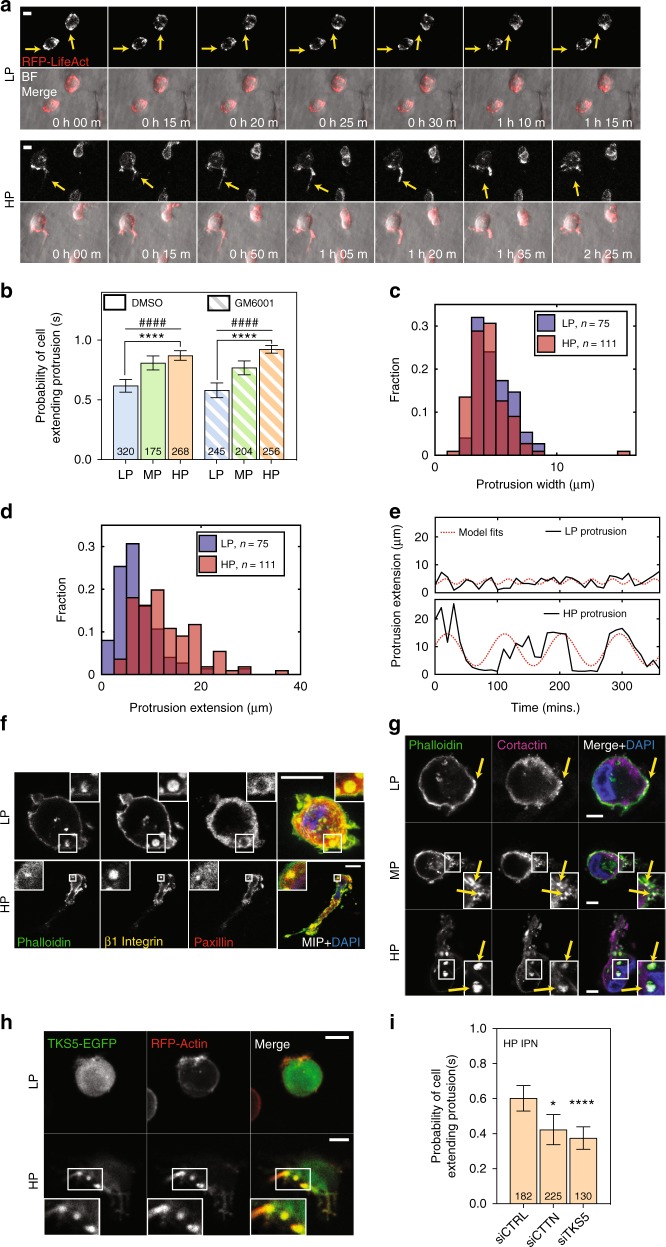
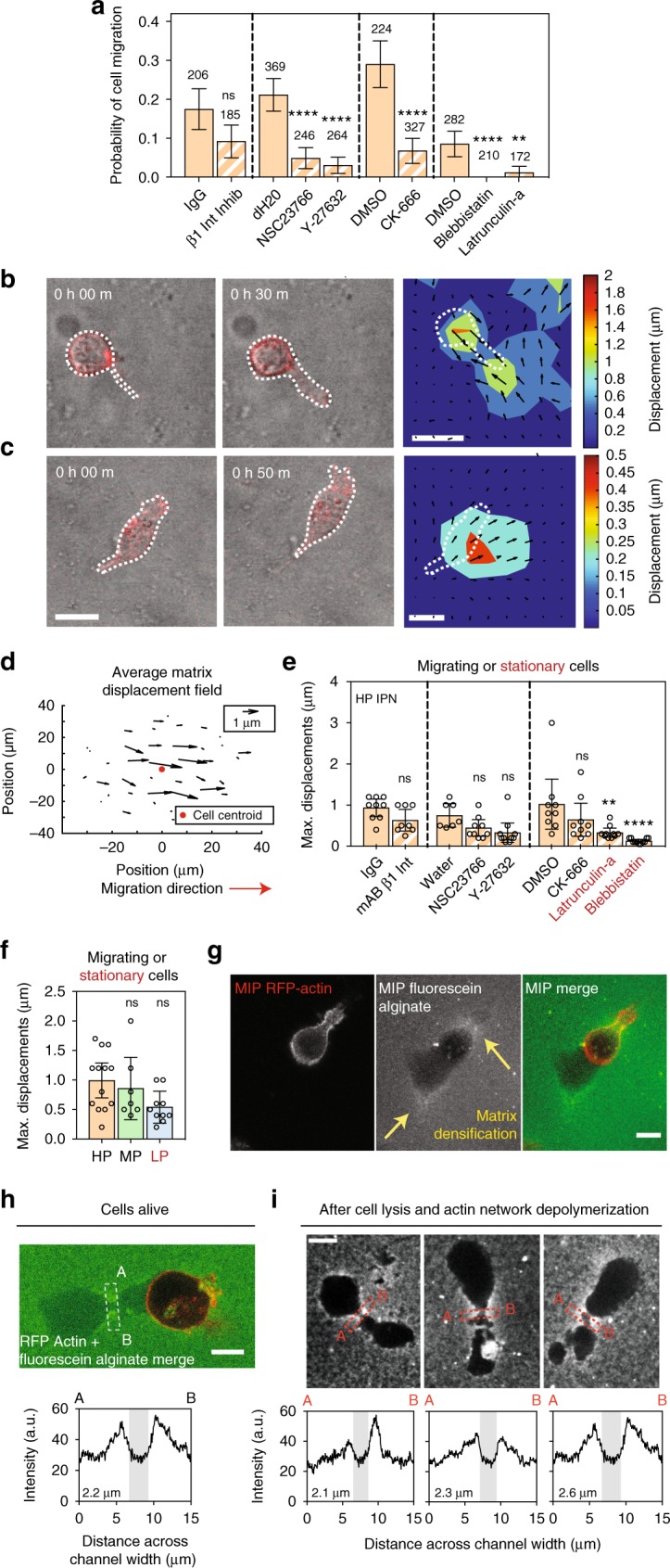
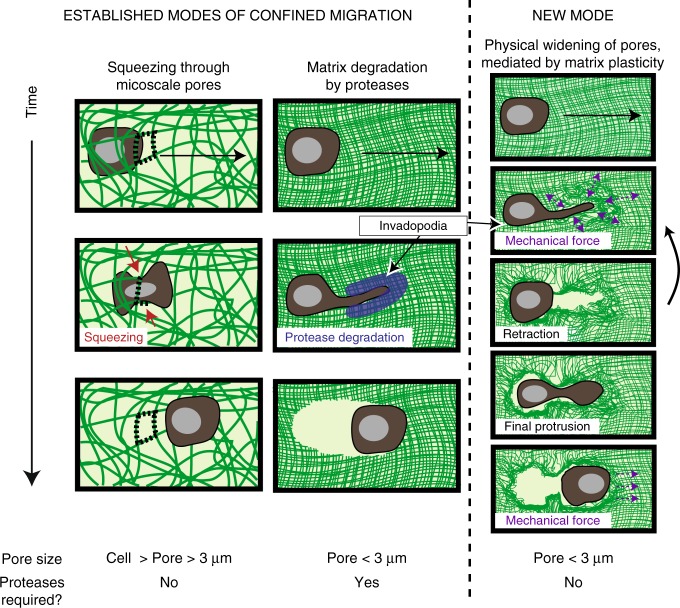
Studies of cancer cell migration have found two modes: one that is protease-independent, requiring micron-sized pores or channels for cells to squeeze through, and one that is protease-dependent, relevant for confining nanoporous matrices such as basement membranes (BMs). However, many extracellular matrices exhibit viscoelasticity and mechanical plasticity, irreversibly deforming in response to force, so that pore size may be malleable. Here we report the impact of matrix plasticity on migration. We develop nanoporous and BM ligand-presenting interpenetrating network (IPN) hydrogels in which plasticity could be modulated independent of stiffness. Strikingly, cells in high plasticity IPNs carry out protease-independent migration through the IPNs. Mechanistically, cells in high plasticity IPNs extend invadopodia protrusions to mechanically and plastically open up micron-sized channels and then migrate through them. These findings uncover a new mode of protease-independent migration, in which cells can migrate through confining matrix if it exhibits sufficient mechanical plasticity.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- RSG-16-028-01/American Cancer Society (American Cancer Society, Inc.)/International
- NNF15OC0015218/Novo Nordisk UK Research Foundation/International
- K99 CA201304/CA/NCI NIH HHS/United States
- CA205262/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)/International
- R21 CA205262/CA/NCI NIH HHS/United States
- R01 GM129098/GM/NIGMS NIH HHS/United States
- R37 CA214136/CA/NCI NIH HHS/United States
- K99CA201304/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)/International
- R37 CA214136/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)/International
LinkOut - more resources
Full Text Sources
Medical
