

Direct protein-lipid interactions shape the conformational landscape of secondary transporters
- PMID: 30297844
- PMCID: PMC6175955
- DOI: 10.1038/s41467-018-06704-1
Direct protein-lipid interactions shape the conformational landscape of secondary transporters
Abstract
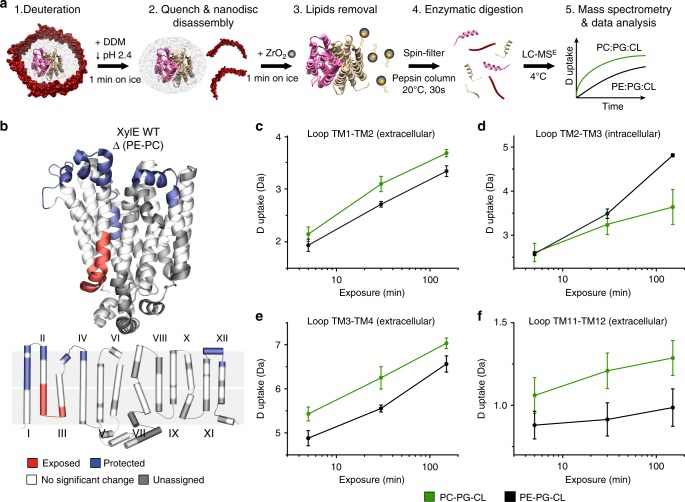
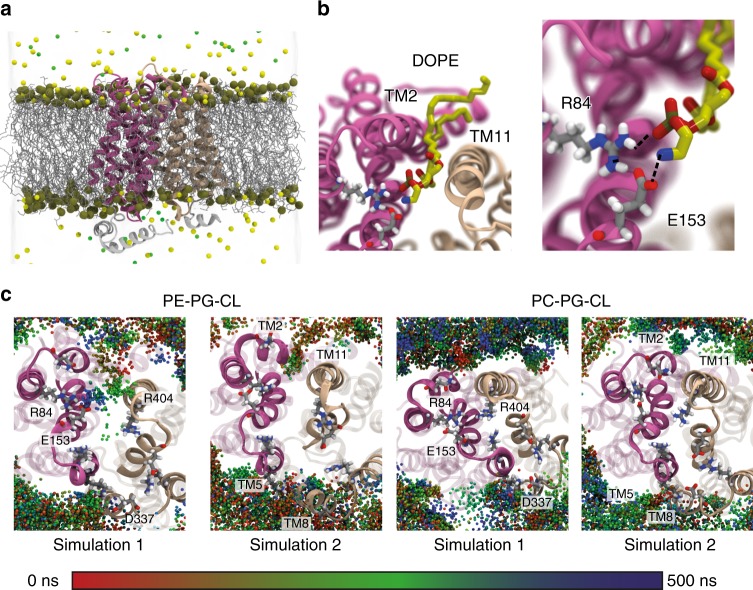
Secondary transporters undergo structural rearrangements to catalyze substrate translocation across the cell membrane - yet how such conformational changes happen within a lipid environment remains poorly understood. Here, we combine hydrogen-deuterium exchange mass spectrometry (HDX-MS) with molecular dynamics (MD) simulations to understand how lipids regulate the conformational dynamics of secondary transporters at the molecular level. Using the homologous transporters XylE, LacY and GlpT from Escherichia coli as model systems, we discover that conserved networks of charged residues act as molecular switches that drive the conformational transition between different states. We reveal that these molecular switches are regulated by interactions with surrounding phospholipids and show that phosphatidylethanolamine interferes with the formation of the conserved networks and favors an inward-facing state. Overall, this work provides insights into the importance of lipids in shaping the conformational landscape of an important class of transporters.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Marger MD, Saier MH., Jr. A major superfamily of transmembrane facilitators that catalyse uniport, symport and antiport. Trends Biochem. Sci. 1993;18:13–20. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01-GM123455/U.S. Department of Health & Human Services | National Institutes of Health (NIH)/International
- 109854/Z/15/Z/WT_/Wellcome Trust/United Kingdom
- MC_PC_15031/MRC_/Medical Research Council/United Kingdom
- R01 GM123455/GM/NIGMS NIH HHS/United States
- U54 GM087519/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
