

MCT2 mediates concentration-dependent inhibition of glutamine metabolism by MOG
- PMID: 30297875
- PMCID: PMC6298574
- DOI: 10.1038/s41589-018-0136-y
MCT2 mediates concentration-dependent inhibition of glutamine metabolism by MOG
Erratum in
-
Author Correction: MCT2 mediates concentration-dependent inhibition of glutamine metabolism by MOG.Nat Chem Biol. 2019 Dec;15(12):1249. doi: 10.1038/s41589-019-0409-0. Nat Chem Biol. 2019. PMID: 31645751
Abstract
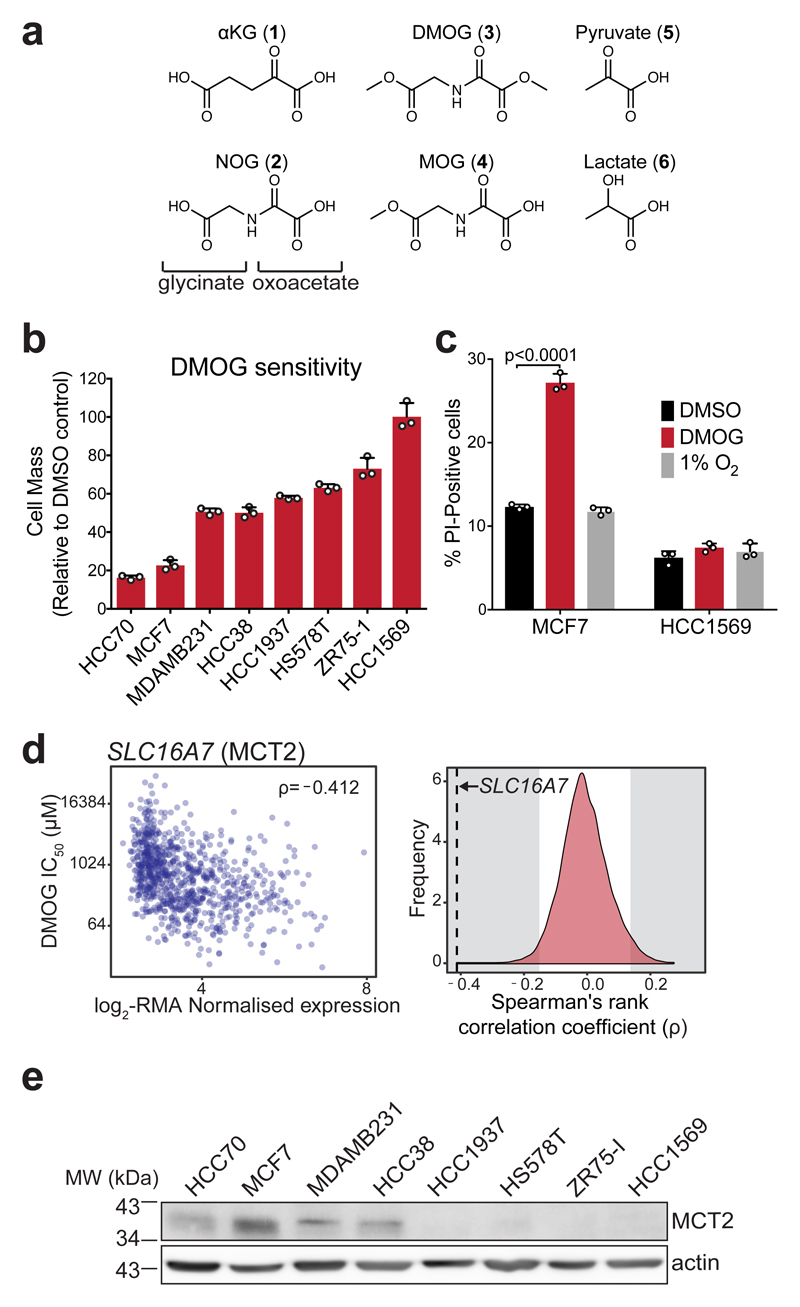
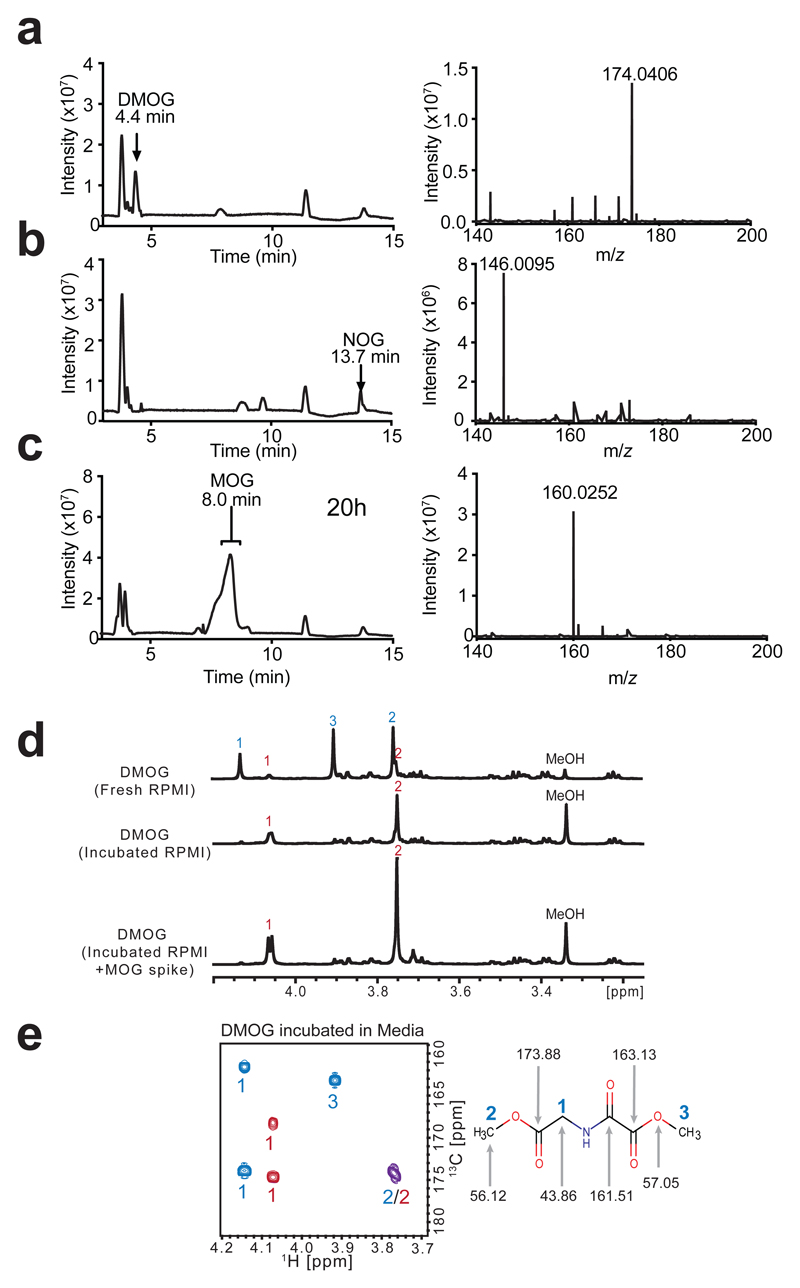
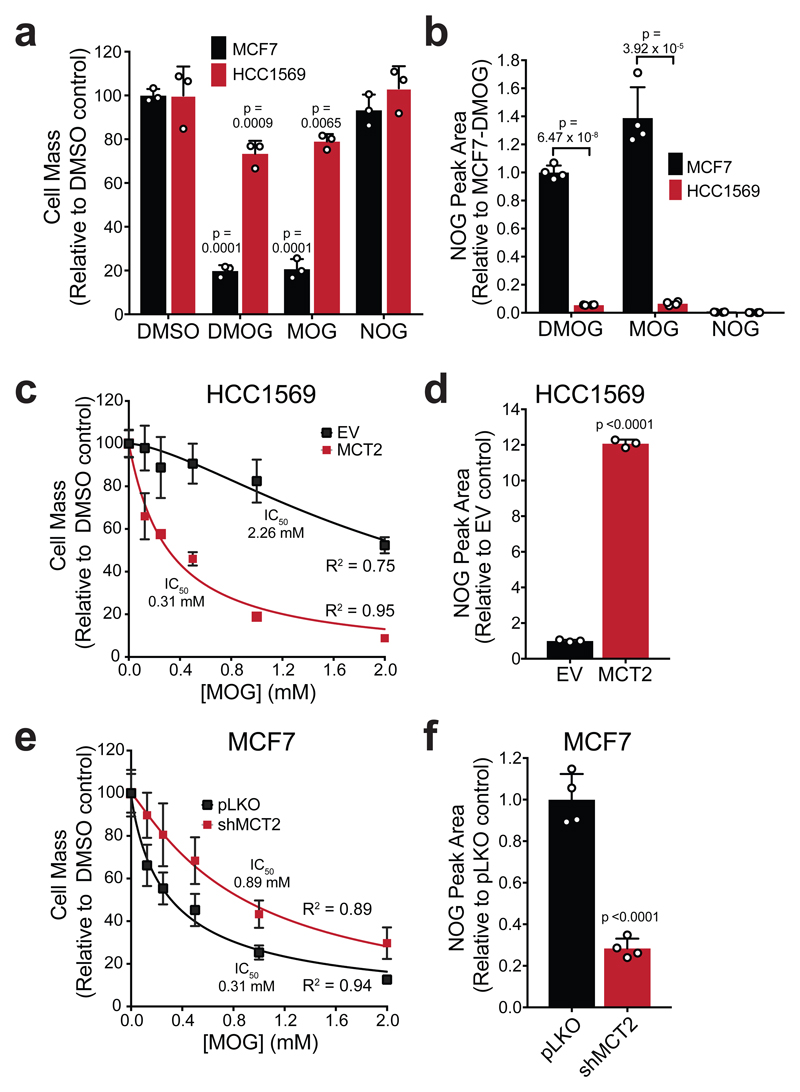
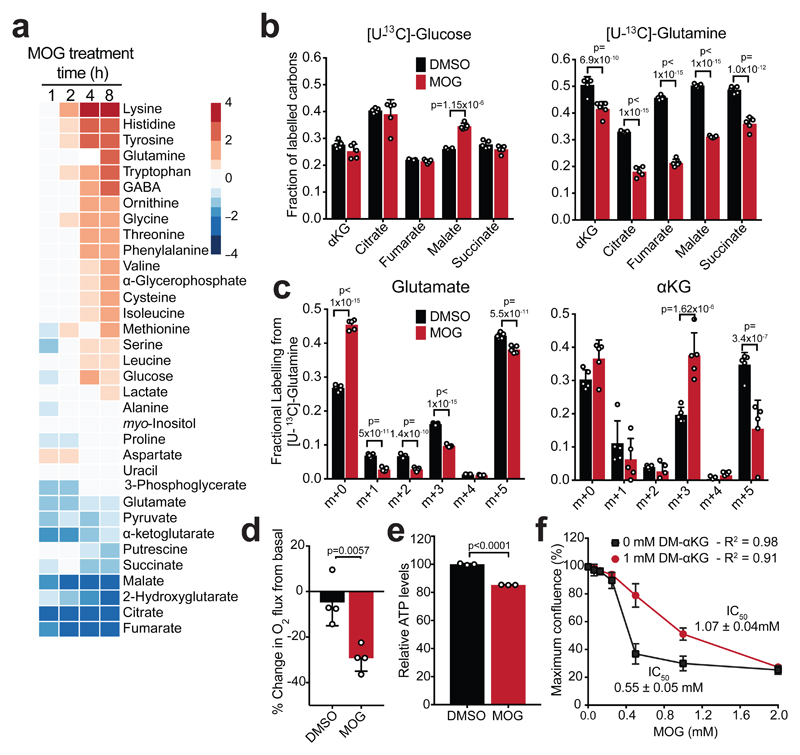
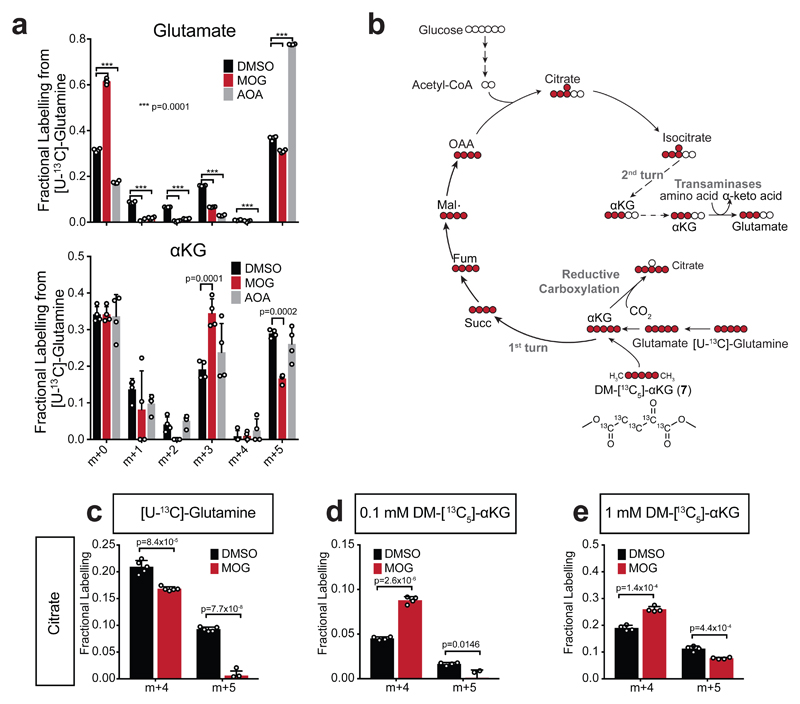
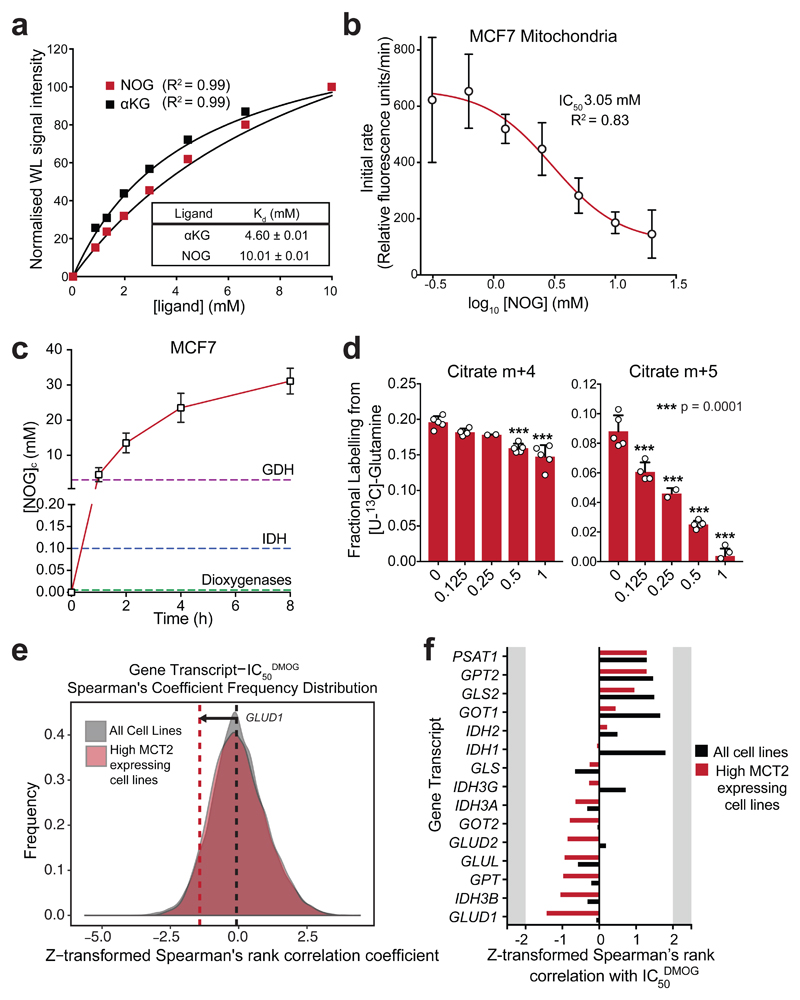
α-Ketoglutarate (αKG) is a key node in many important metabolic pathways. The αKG analog N-oxalylglycine (NOG) and its cell-permeable prodrug dimethyloxalylglycine (DMOG) are extensively used to inhibit αKG-dependent dioxygenases. However, whether NOG interference with other αKG-dependent processes contributes to its mode of action remains poorly understood. Here we show that, in aqueous solutions, DMOG is rapidly hydrolyzed, yielding methyloxalylglycine (MOG). MOG elicits cytotoxicity in a manner that depends on its transport by monocarboxylate transporter 2 (MCT2) and is associated with decreased glutamine-derived tricarboxylic acid-cycle flux, suppressed mitochondrial respiration and decreased ATP production. MCT2-facilitated entry of MOG into cells leads to sufficiently high concentrations of NOG to inhibit multiple enzymes in glutamine metabolism, including glutamate dehydrogenase. These findings reveal that MCT2 dictates the mode of action of NOG by determining its intracellular concentration and have important implications for the use of (D)MOG in studying αKG-dependent signaling and metabolism.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
