

Late-stage tumors induce anemia and immunosuppressive extramedullary erythroid progenitor cells
- PMID: 30297899
- PMCID: PMC6211844
- DOI: 10.1038/s41591-018-0205-5
Late-stage tumors induce anemia and immunosuppressive extramedullary erythroid progenitor cells
Abstract
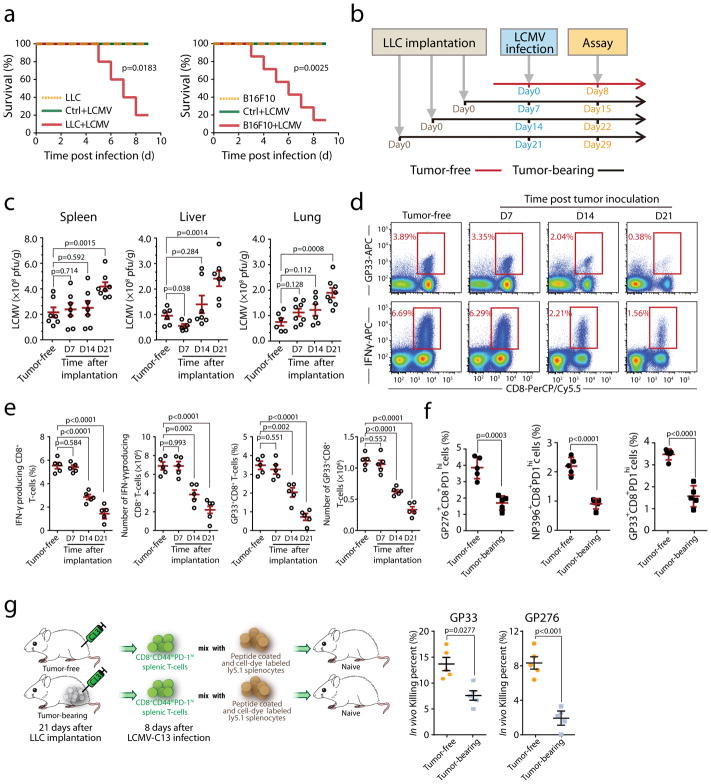
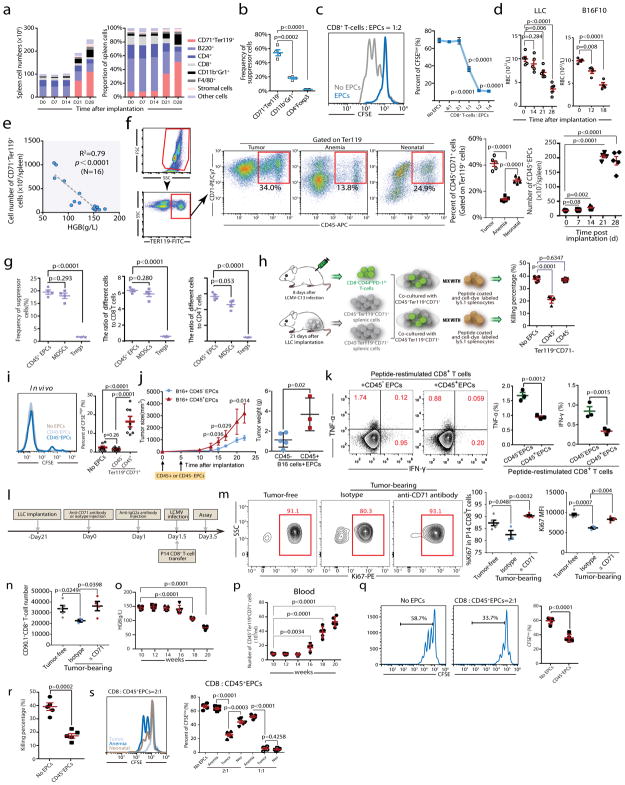
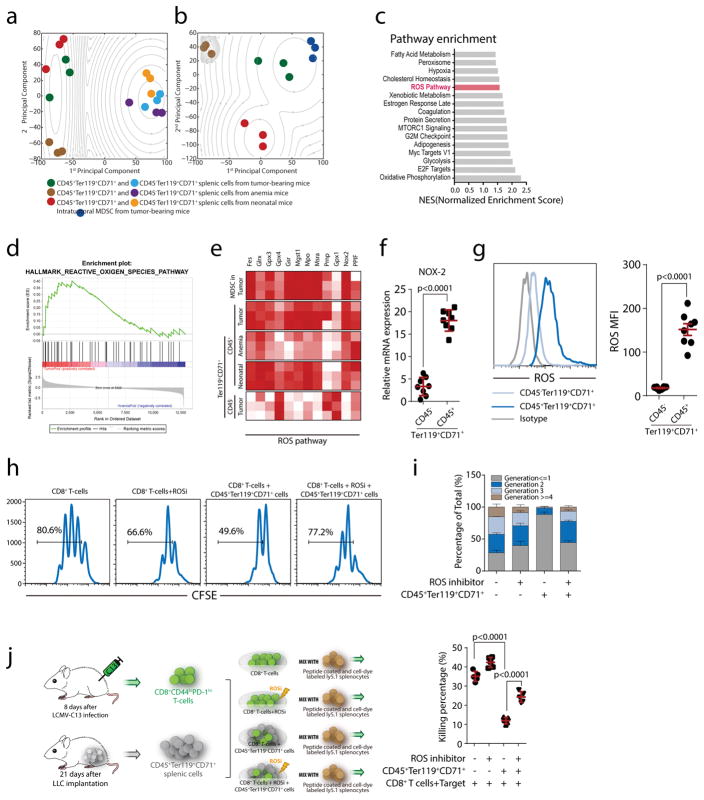
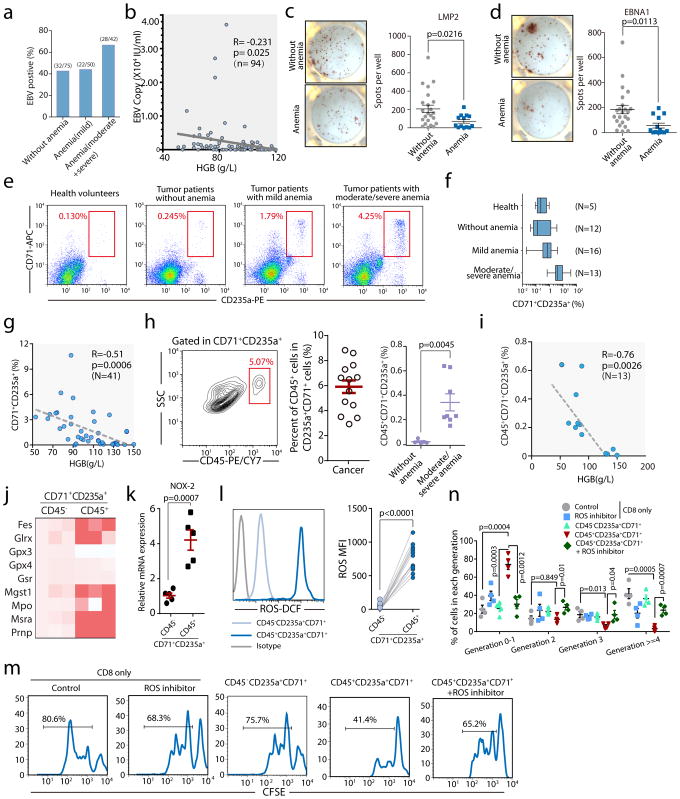
Impaired immunity in patients with late-stage cancer is not limited to antitumor responses, as demonstrated by poor vaccination protection and high susceptibility to infection1-3. This has been largely attributed to chemotherapy-induced impairment of innate immunity, such as neutropenia2, whereas systemic effects of tumors on hematopoiesis and adoptive immunity remain incompletely understood. Here we observed anemia associated with severe deficiency of CD8+ T cell responses against pathogens in treatment-naive mice bearing large tumors. Specifically, we identify CD45+ erythroid progenitor cells (CD71+TER119+; EPCs) as robust immunosuppressors. CD45+ EPCs, induced by tumor growth-associated extramedullary hematopoiesis, accumulate in the spleen to become a major population, outnumbering regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs). The CD45+ EPC transcriptome closely resembles that of MDSCs, and, like MDSCs, reactive oxygen species production is a major mechanism underlying CD45+ EPC-mediated immunosuppression. Similarly, an immunosuppressive CD45+ EPC population was detected in patients with cancer who have anemia. These findings identify a major population of immunosuppressive cells that likely contributes to the impaired T cell responses commonly observed in patients with advanced cancer.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Baden LR, et al. Prevention and Treatment of Cancer-Related Infections. Journal of the National Comprehensive Cancer Network. 2012;10:1412–1445. - PubMed
-
- Vento S, Cainelli F, Ternesgen Z. Lung infections after cancer chemotherapy. Lancet Oncology. 2008;9:982–992. - PubMed
-
- van der Burg SH, Arens R, Ossendorp F, van Hall T, Melief AJM. Vaccines for established cancer: overcoming the challenges posed by immune evasion. Nature Reviews Cancer. 2016;16:219–233. - PubMed
-
- Bodey GP. Infection in cancer patients. A continuing association. The American journal of medicine. 1986;81:11–26. - PubMed
-
- Kosmidis CI, Chandrasekar PH. Management of gram-positive bacterial infections in patients with cancer. Leukemia & Lymphoma. 2012;53:8–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
