

Structure-guided combination therapy to potently improve the function of mutant CFTRs
- PMID: 30297908
- PMCID: PMC6301090
- DOI: 10.1038/s41591-018-0200-x
Structure-guided combination therapy to potently improve the function of mutant CFTRs
Abstract
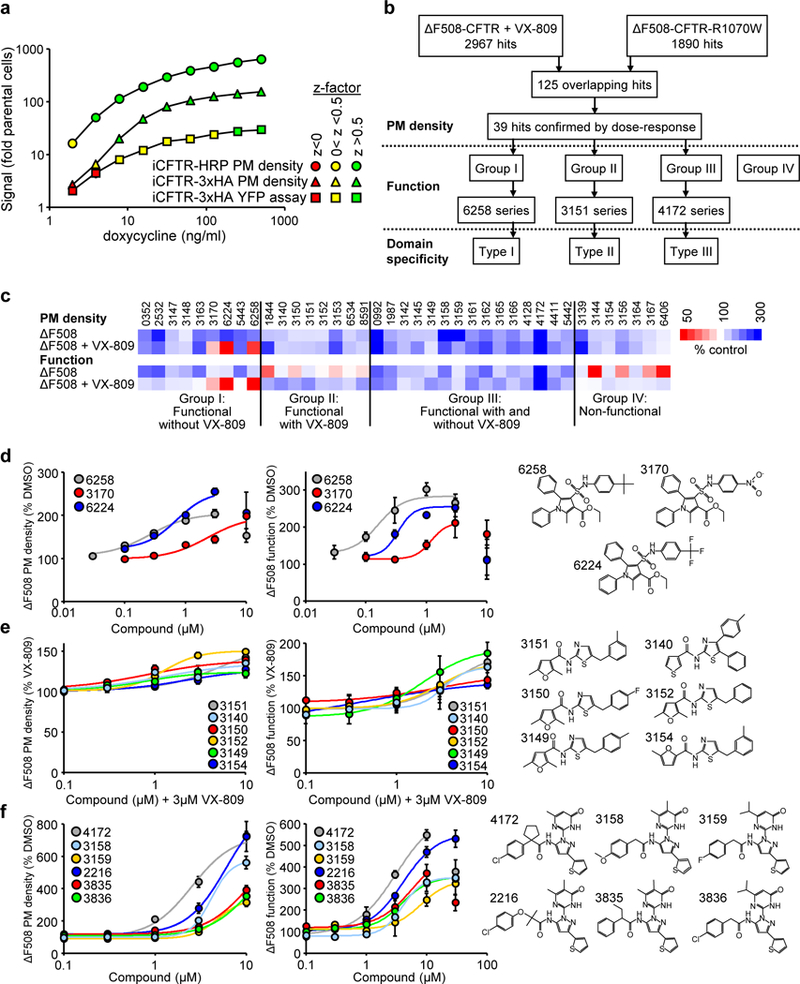
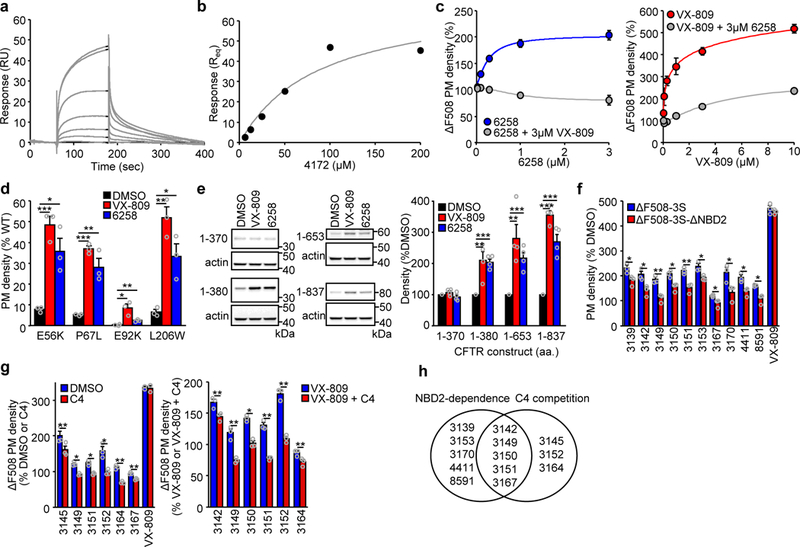
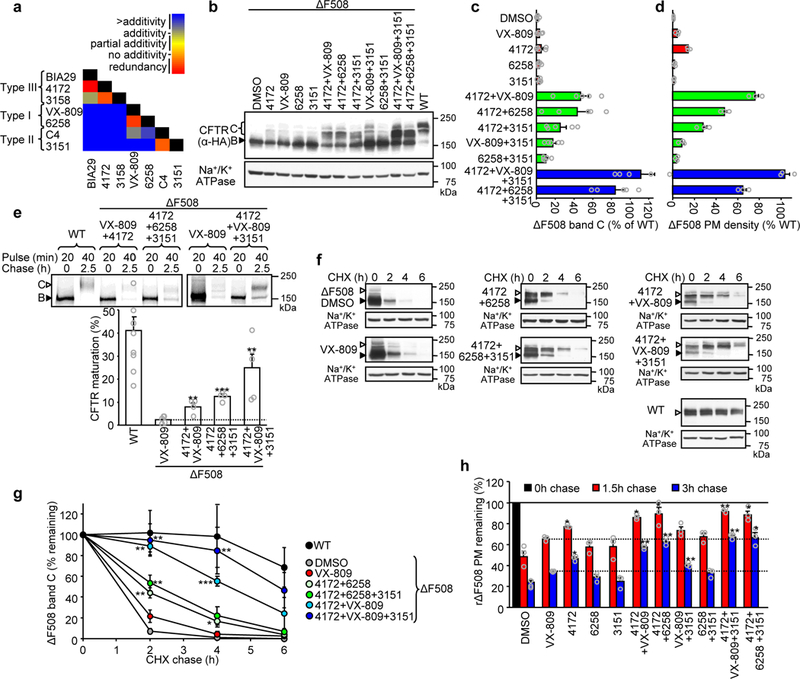
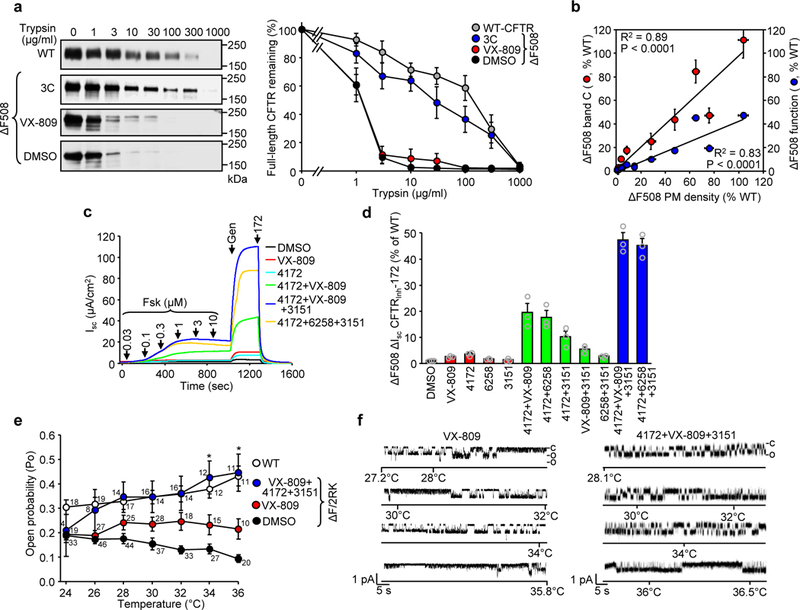
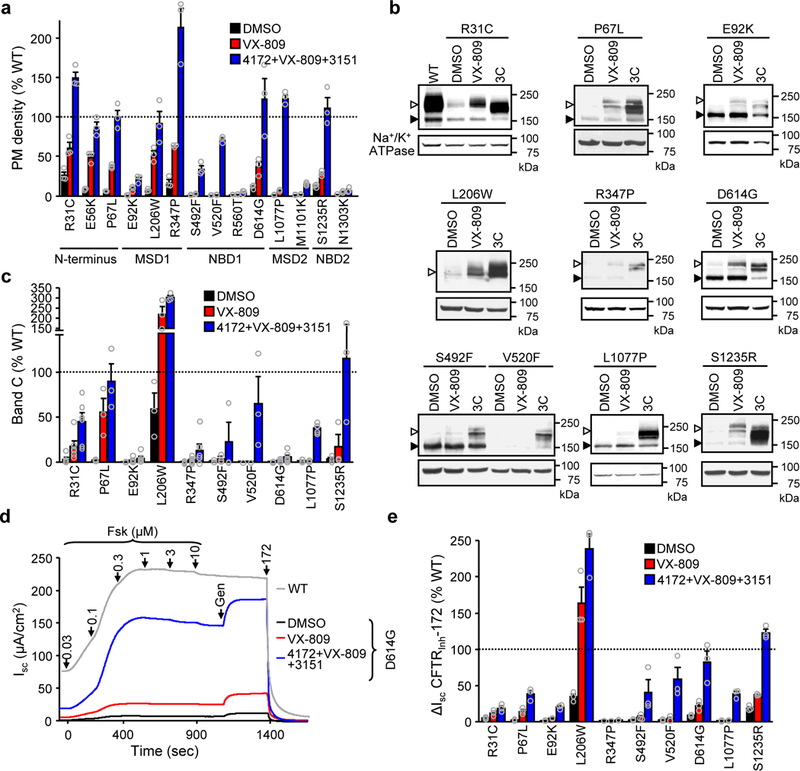
Available corrector drugs are unable to effectively rescue the folding defects of CFTR-ΔF508 (or CFTR-F508del), the most common disease-causing mutation of the cystic fibrosis transmembrane conductance regulator, a plasma membrane (PM) anion channel, and thus to substantially ameliorate clinical phenotypes of cystic fibrosis (CF). To overcome the corrector efficacy ceiling, here we show that compounds targeting distinct structural defects of CFTR can synergistically rescue mutant expression and function at the PM. High-throughput cell-based screens and mechanistic analysis identified three small-molecule series that target defects at nucleotide-binding domain (NBD1), NBD2 and their membrane-spanning domain (MSD) interfaces. Although individually these compounds marginally improve ΔF508-CFTR folding efficiency, function and stability, their combinations lead to ~50-100% of wild-type-level correction in immortalized and primary human airway epithelia and in mouse nasal epithelia. Likewise, corrector combinations were effective against rare missense mutations in various CFTR domains, probably acting via structural allostery, suggesting a mechanistic framework for their broad application.
Conflict of interest statement
Competing financial interests
C.L., W.L., K.M., S.G., P.A.M., F.J.K., E.A., A.J.O., P.M. and W.G.B. are employees of the Genomics Institute of the Novartis Research Foundation. I.S.-G. has been principal investigator in Vertex initiated clinical trials, received a Vertex Pharmaceuticals Innovation Award and served as a scientific advisory board member for Vertex Pharmaceuticals. G.L.L. is a member of the Scientific Advisory Board of Proteostasis Therapeutics Inc. All other authors declare no competing financial or non-financial interests.
Figures

References
-
- Liu F, Zhang Z, Csanady L, Gadsby DC & Chen J Molecular Structure of the Human CFTR Ion Channel. Cell 169, 85–95 e88 (2017). - PubMed
Methods-only References
-
- Neuberger T, Burton B, Clark H & Van Goor F Use of primary cultures of human bronchial epithelial cells isolated from cystic fibrosis patients for the pre-clinical testing of CFTR modulators. Methods Mol Biol 741, 39–54 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
