

Integrative transcriptome analyses of the aging brain implicate altered splicing in Alzheimer's disease susceptibility
- PMID: 30297968
- PMCID: PMC6354244
- DOI: 10.1038/s41588-018-0238-1
Integrative transcriptome analyses of the aging brain implicate altered splicing in Alzheimer's disease susceptibility
Abstract
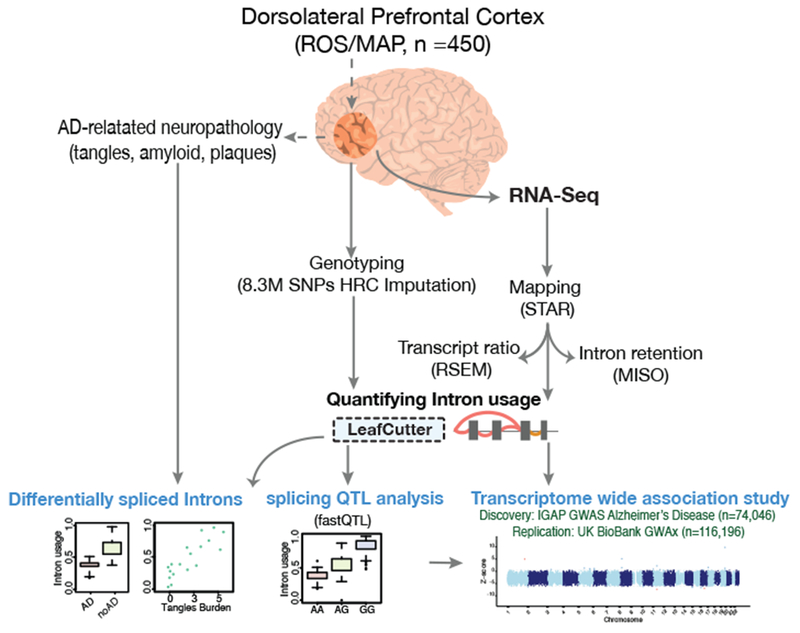
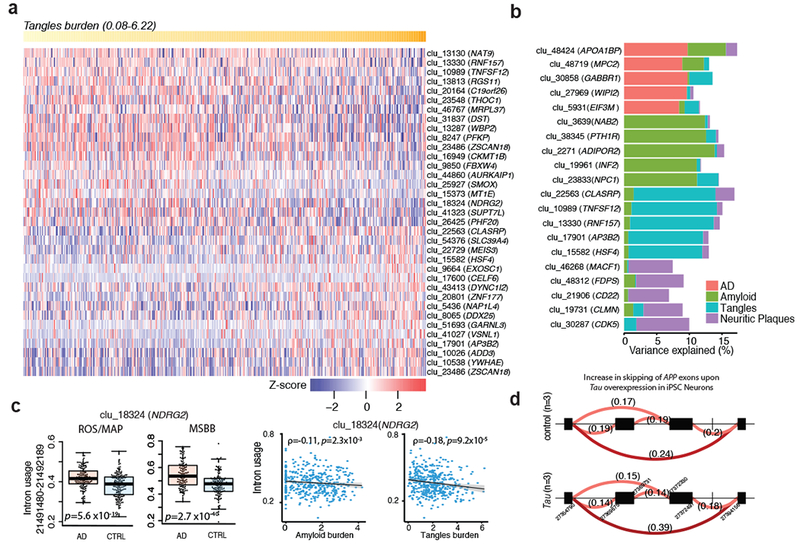
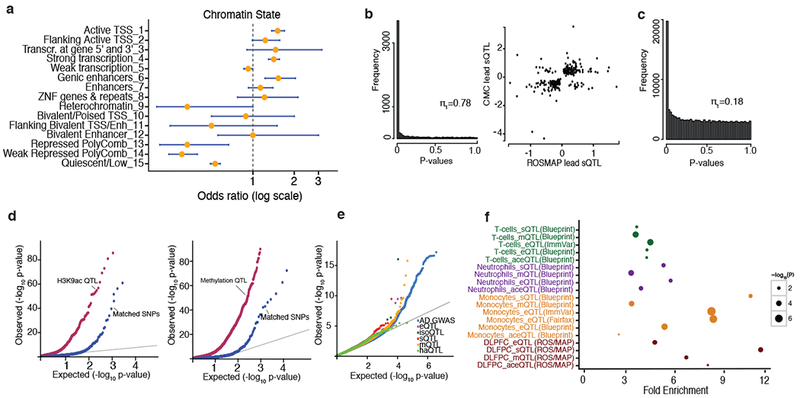
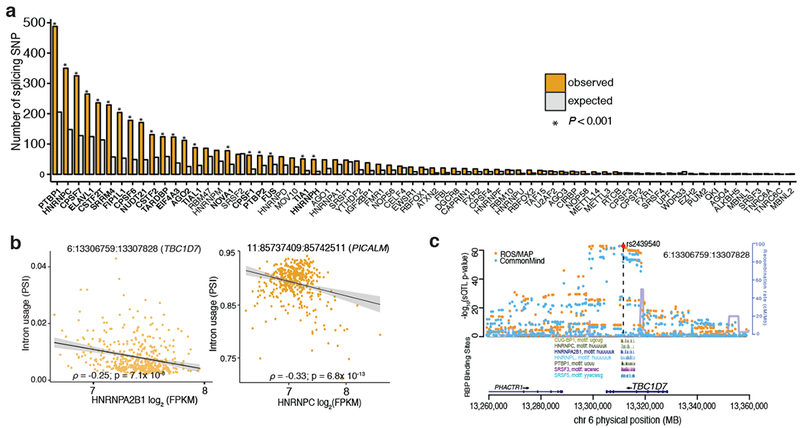
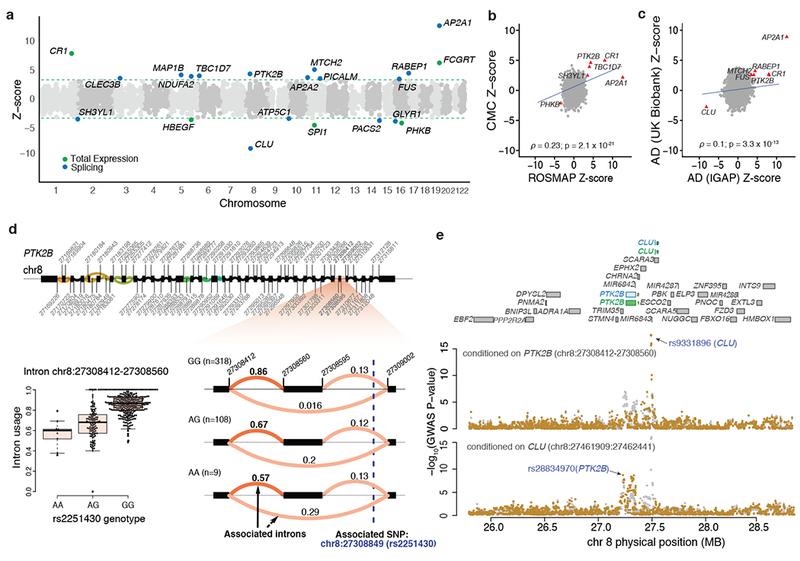
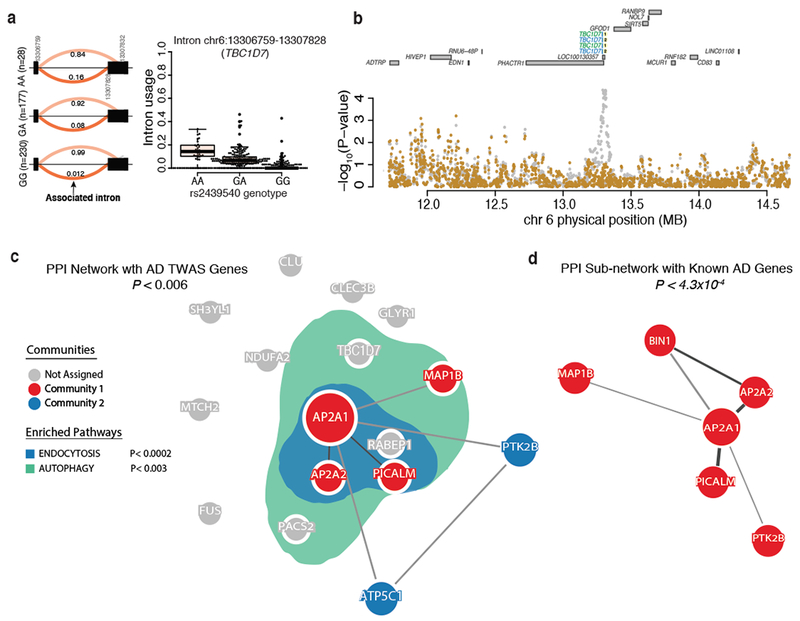
Here we use deep sequencing to identify sources of variation in mRNA splicing in the dorsolateral prefrontal cortex (DLPFC) of 450 subjects from two aging cohorts. Hundreds of aberrant pre-mRNA splicing events are reproducibly associated with Alzheimer's disease. We also generate a catalog of splicing quantitative trait loci (sQTL) effects: splicing of 3,006 genes is influenced by genetic variation. We report that altered splicing is the mechanism for the effects of the PICALM, CLU and PTK2B susceptibility alleles. Furthermore, we performed a transcriptome-wide association study and identified 21 genes with significant associations with Alzheimer's disease, many of which are found in known loci, whereas 8 are in novel loci. These results highlight the convergence of old and new genes associated with Alzheimer's disease in autophagy-lysosomal-related pathways. Overall, this study of the transcriptome of the aging brain provides evidence that dysregulation of mRNA splicing is a feature of Alzheimer's disease and is, in some cases, genetically driven.
Conflict of interest statement
Competing financial interests
The authors declare no competing financial interests.
Figures
Comment in
-
Altered splicing in Alzheimer transcriptomes.Nat Rev Genet. 2018 Dec;19(12):738-739. doi: 10.1038/s41576-018-0064-4. Nat Rev Genet. 2018. PMID: 30341439 No abstract available.
References
-
- Kornblihtt AR et al. Alternative splicing: a pivotal step between eukaryotic transcription and translation. Nat Rev Mol Cell Biol 14, 153–65 (2013). - PubMed
-
- Barbosa-Morais NL et al. The evolutionary landscape of alternative splicing in vertebrate species. Science 338, 1587–93 (2012). - PubMed
-
- Wang GS & Cooper TA Splicing in disease: disruption of the splicing code and the decoding machinery. Nat Rev Genet 8, 749–61 (2007). - PubMed
-
- Dredge BK, Polydorides AD & Darnell RB The splice of life: alternative splicing and neurological disease. Nat Rev Neurosci 2, 43–50 (2001). - PubMed
Publication types
MeSH terms
Grants and funding
- U01 AG046152/AG/NIA NIH HHS/United States
- R01 AG054005/AG/NIA NIH HHS/United States
- R37 MH057881/MH/NIMH NIH HHS/United States
- R01 AG017917/AG/NIA NIH HHS/United States
- S10 OD018522/OD/NIH HHS/United States
- P30 AG010161/AG/NIA NIH HHS/United States
- P50 MH084053/MH/NIMH NIH HHS/United States
- P50 MH066392/MH/NIMH NIH HHS/United States
- R01 MH080405/MH/NIMH NIH HHS/United States
- R01 MH075916/MH/NIMH NIH HHS/United States
- R56 AG055824/AG/NIA NIH HHS/United States
- R01 MH097276/MH/NIMH NIH HHS/United States
- P01 AG002219/AG/NIA NIH HHS/United States
- R01 MH093725/MH/NIMH NIH HHS/United States
- P50 AG005138/AG/NIA NIH HHS/United States
- R01 AG036836/AG/NIA NIH HHS/United States
- R01 MH085542/MH/NIMH NIH HHS/United States
- R01 AG015819/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
