

Biochemical characterization of the minimal domains of an iterative eukaryotic polyketide synthase
- PMID: 30300504
- PMCID: PMC6334511
- DOI: 10.1111/febs.14675
Biochemical characterization of the minimal domains of an iterative eukaryotic polyketide synthase
Abstract
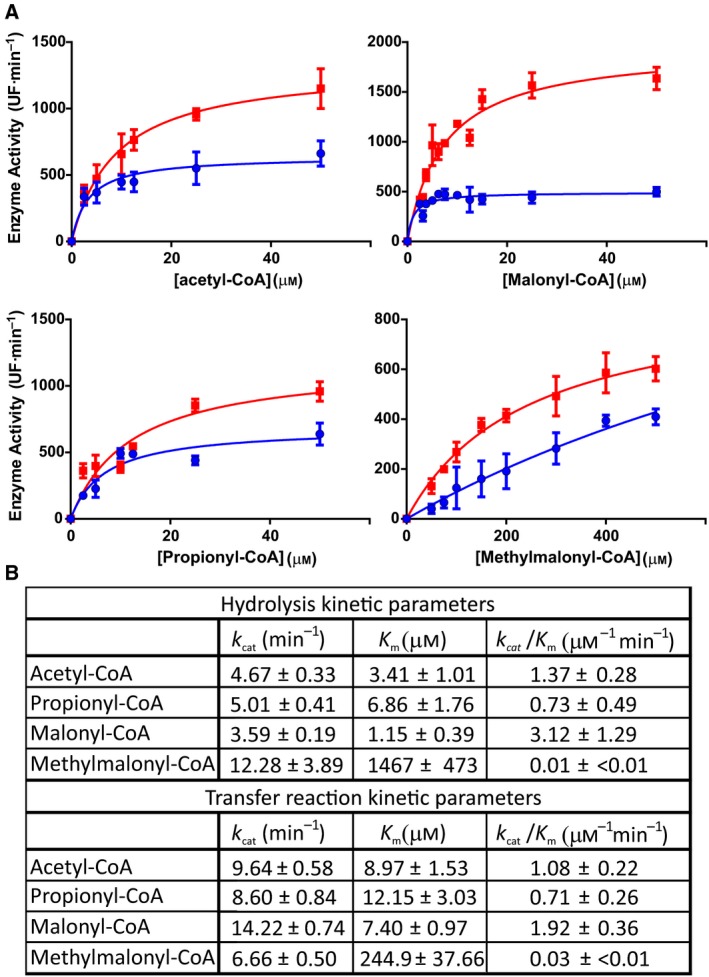
Iterative type I polyketide synthases (PKS) are megaenzymes essential to the biosynthesis of an enormously diverse array of bioactive natural products. Each PKS contains minimally three functional domains, β-ketosynthase (KS), acyltransferase (AT), and acyl carrier protein (ACP), and a subset of reducing domains such as ketoreductase (KR), dehydratase (DH), and enoylreductase (ER). The substrate selection, condensation reactions, and β-keto processing of the polyketide growing chain are highly controlled in a programmed manner. However, the structural features and mechanistic rules that orchestrate the iterative cycles, processing domains functionality, and chain termination in this kind of megaenzymes are often poorly understood. Here, we present a biochemical and functional characterization of the KS and the AT domains of a PKS from the mallard duck Anas platyrhynchos (ApPKS). ApPKS belongs to an animal PKS family phylogenetically more related to bacterial PKS than to metazoan fatty acid synthases. Through the dissection of the ApPKS enzyme into mono- to didomain fragments and its reconstitution in vitro, we determined its substrate specificity toward different starters and extender units. ApPKS AT domain can effectively transfer acetyl-CoA and malonyl-CoA to the ApPKS ACP stand-alone domain. Furthermore, the KS and KR domains, in the presence of Escherichia coli ACP, acetyl-CoA, and malonyl-CoA, showed the ability to catalyze the chain elongation and the β-keto reduction steps necessary to yield a 3-hydroxybutyryl-ACP derivate. These results provide new insights into the catalytic efficiency and specificity of this uncharacterized family of PKSs.
Keywords: PKS biochemistry; domain deconstruction; iterative PKS; substrate specificity.
© 2018 The Authors. The FEBS Journal published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Figures






Similar articles
-
Reconstituting modular activity from separated domains of 6-deoxyerythronolide B synthase.Biochemistry. 2004 Nov 9;43(44):13892-8. doi: 10.1021/bi048418n. Biochemistry. 2004. PMID: 15518537
-
Extender unit and acyl carrier protein specificity of ketosynthase domains of the 6-deoxyerythronolide B synthase.J Am Chem Soc. 2006 Mar 8;128(9):3067-74. doi: 10.1021/ja058093d. J Am Chem Soc. 2006. PMID: 16506788 Free PMC article.
-
Engineering the acyltransferase domain of epothilone polyketide synthase to alter the substrate specificity.Microb Cell Fact. 2021 Apr 21;20(1):86. doi: 10.1186/s12934-021-01578-3. Microb Cell Fact. 2021. PMID: 33882930 Free PMC article.
-
The stereochemistry of complex polyketide biosynthesis by modular polyketide synthases.Molecules. 2011 Jul 20;16(7):6092-115. doi: 10.3390/molecules16076092. Molecules. 2011. PMID: 21775938 Free PMC article. Review.
-
The architectures of iterative type I PKS and FAS.Nat Prod Rep. 2018 Oct 17;35(10):1046-1069. doi: 10.1039/c8np00039e. Nat Prod Rep. 2018. PMID: 30137093 Free PMC article. Review.
Cited by
-
Chromosome-level genome assembly of the sacoglossan sea slug Elysia timida (Risso, 1818).BMC Genomics. 2024 Oct 7;25(1):941. doi: 10.1186/s12864-024-10829-7. BMC Genomics. 2024. PMID: 39375624 Free PMC article.
-
The mud-dwelling clam Meretrix petechialis secretes endogenously synthesized erythromycin.Proc Natl Acad Sci U S A. 2022 Dec 6;119(49):e2214150119. doi: 10.1073/pnas.2214150119. Epub 2022 Nov 28. Proc Natl Acad Sci U S A. 2022. PMID: 36442100 Free PMC article.
-
Diverse host-associated fungal systems as a dynamic source of novel bioactive anthraquinones in drug discovery: Current status and future perspectives.J Adv Res. 2022 Jul;39:257-273. doi: 10.1016/j.jare.2021.11.007. Epub 2021 Nov 22. J Adv Res. 2022. PMID: 35660073 Free PMC article. Review.
-
Genome Mining of Pseudanabaena galeata CCNP1313 Indicates a New Scope in the Search for Antiproliferative and Antiviral Agents.Microorganisms. 2024 Aug 9;12(8):1628. doi: 10.3390/microorganisms12081628. Microorganisms. 2024. PMID: 39203471 Free PMC article.
-
Animal biosynthesis of complex polyketides in a photosynthetic partnership.Nat Commun. 2020 Jun 8;11(1):2882. doi: 10.1038/s41467-020-16376-5. Nat Commun. 2020. PMID: 32513940 Free PMC article.
References
-
- Staunton J & Weissman KJ (2001) Polyketide biosynthesis: a millennium review. Nat Prod Rep 18, 380–416. - PubMed
-
- Gokhale RS, Saxena P, Chopra T & Mohanty D (2007) Versatile polyketide enzymatic machinery for the biosynthesis of complex mycobacterial lipids. Nat Prod Rep 24, 267–277. - PubMed
-
- Medema MH, Blin K, Cimermancic P, De Jager V, Zakrzewski P, Fischbach MA, Weber T, Takano E & Breitling R (2011) AntiSMASH: rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res 39, 339–346. - PMC - PubMed
-
- Rittner A & Grininger M (2014) Modular polyketide synthases (PKSs): a new model fits all? ChemBioChem 15, 2489–2493. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
