

Unilateral, 3D Arm Movement Kinematics Are Encoded in Ipsilateral Human Cortex
- PMID: 30301759
- PMCID: PMC6246886
- DOI: 10.1523/JNEUROSCI.0015-18.2018
Unilateral, 3D Arm Movement Kinematics Are Encoded in Ipsilateral Human Cortex
Abstract
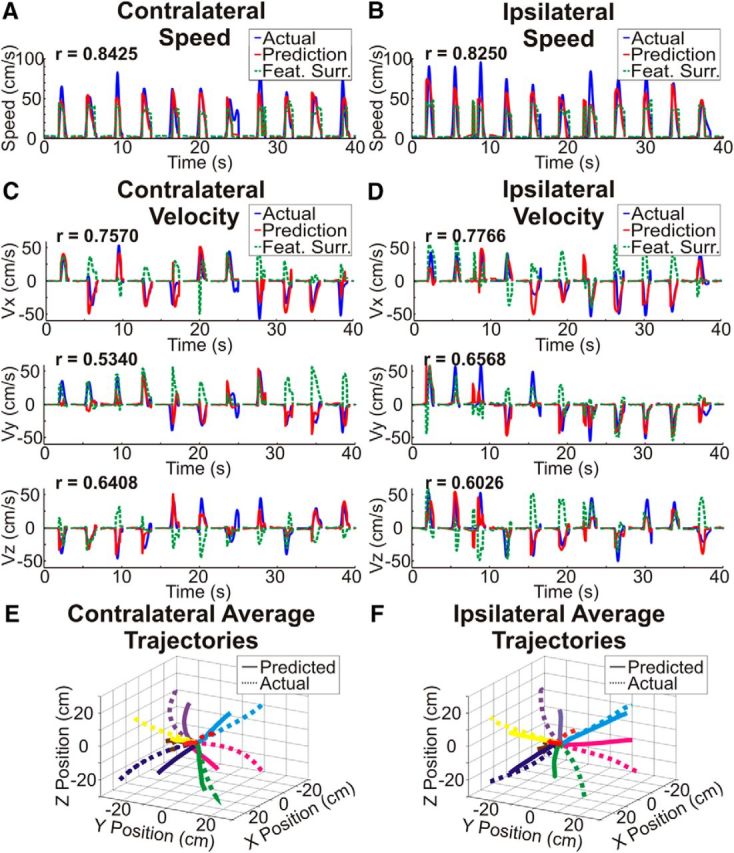
There is increasing evidence that the hemisphere ipsilateral to a moving limb plays a role in planning and executing movements. However, the exact relationship between cortical activity and ipsilateral limb movements is uncertain. We sought to determine whether 3D arm movement kinematics (speed, velocity, and position) could be decoded from cortical signals recorded from the hemisphere ipsilateral to the moving limb. By having invasively monitored patients perform unilateral reaches with each arm, we also compared the encoding of contralateral and ipsilateral limb kinematics from a single cortical hemisphere. In four motor-intact human patients (three male, one female) implanted with electrocorticography electrodes for localization of their epileptic foci, we decoded 3D movement kinematics of both arms with accuracies above chance. Surprisingly, the spatial and spectral encoding of contralateral and ipsilateral limb kinematics was similar, enabling cross-prediction of kinematics between arms. These results clarify our understanding that the ipsilateral hemisphere robustly contributes to motor execution and supports that the information of complex movements is more bihemispherically represented in humans than has been previously understood.SIGNIFICANCE STATEMENT Although limb movements are traditionally understood to be driven by the cortical hemisphere contralateral to a moving limb, movement-related neural activity has also been found in the ipsilateral hemisphere. This study provides the first demonstration that 3D arm movement kinematics can be decoded from human electrocorticographic signals ipsilateral to the moving limb. Surprisingly, the spatial and spectral encoding of contralateral and ipsilateral limb kinematics was similar. The finding that specific kinematics are encoded in the ipsilateral hemisphere demonstrates that the ipsilateral hemisphere contributes to the execution of unilateral limb movements, improving our understanding of motor control. Additionally, the bihemisheric representation of voluntary movements has implications for the development of neuroprosthetic systems for reaching and for neurorehabilitation strategies following cortical injuries.
Keywords: BCI; ECoG; electrocorticography; ipsilateral; reach.
Copyright © 2018 Bundy et al.
Figures







References
-
- Aizawa H, Mushiake H, Inase M, Tanji J (1990) An output zone of the monkey primary motor cortex specialized for bilateral hand movement. Exp Brain Res 82:219–221. - PubMed
-
- Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Stat Soc B Met 57:289–300.
-
- Benjamini Y, Yekutieli D (2001) The control of the false discovery rate in multiple testing under dependency. Ann Stat 29:1165–1188. 10.1214/aos/1013699998 - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources