

Three structurally and functionally distinct β-glucuronidases from the human gut microbe Bacteroides uniformis
- PMID: 30301767
- PMCID: PMC6290157
- DOI: 10.1074/jbc.RA118.005414
Three structurally and functionally distinct β-glucuronidases from the human gut microbe Bacteroides uniformis
Abstract
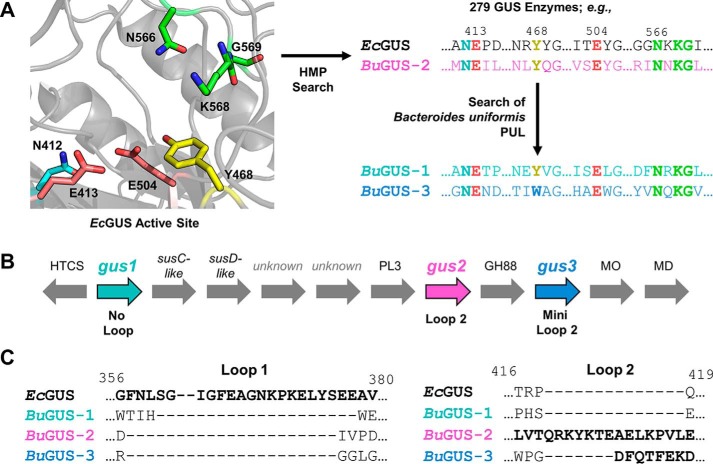
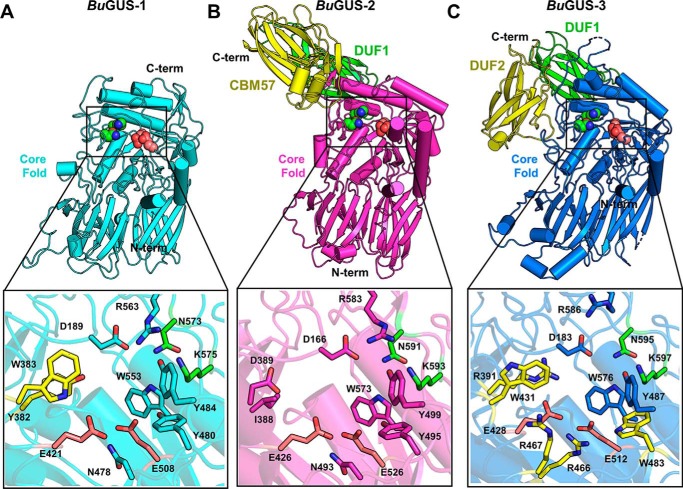
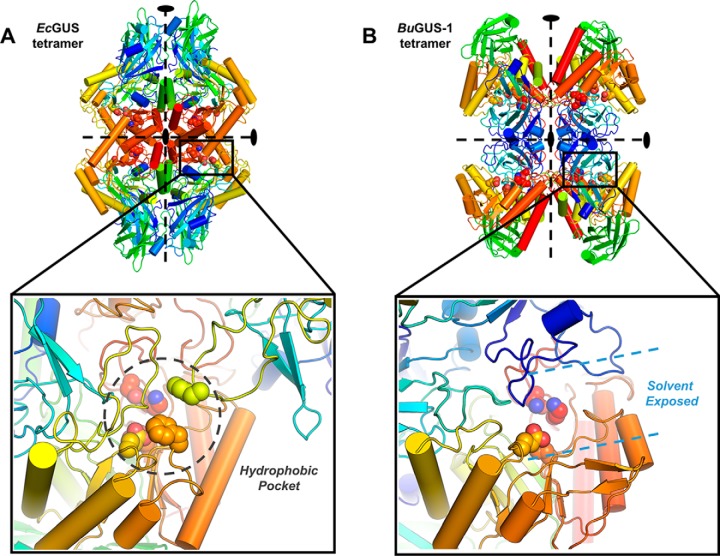
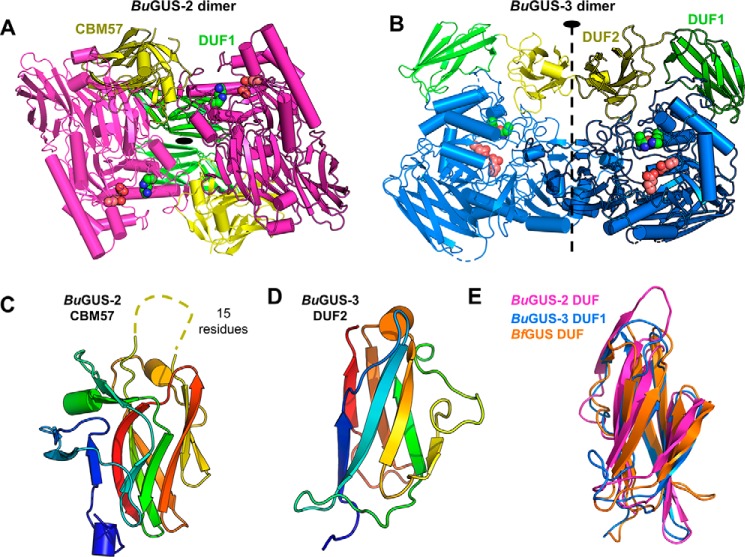
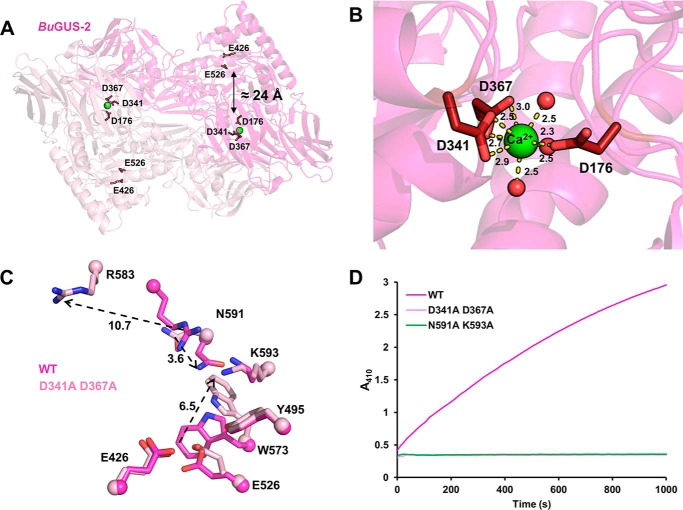
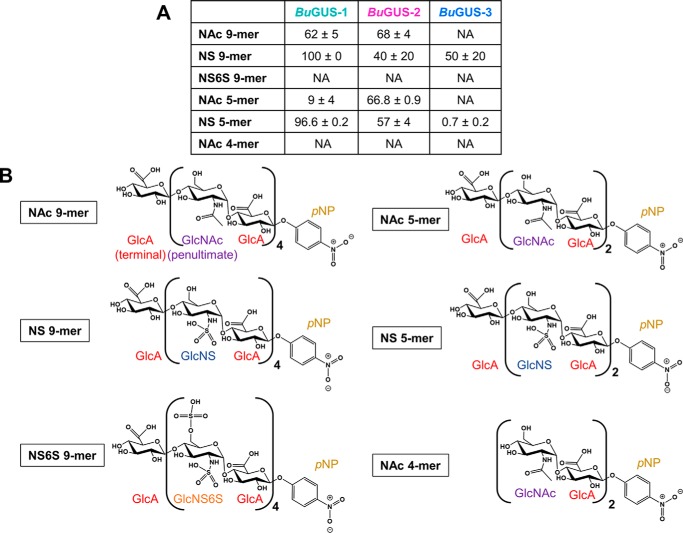
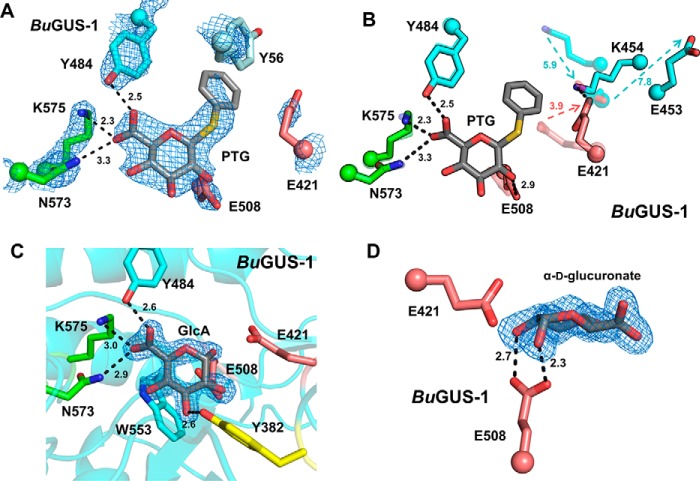
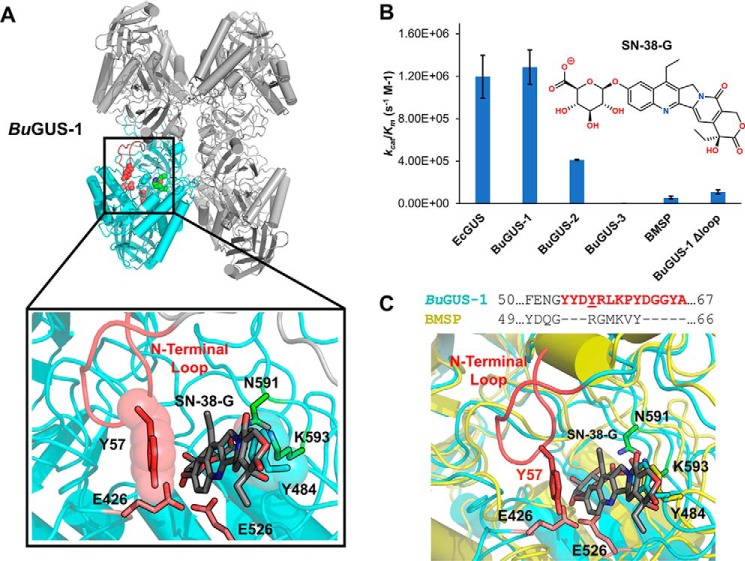
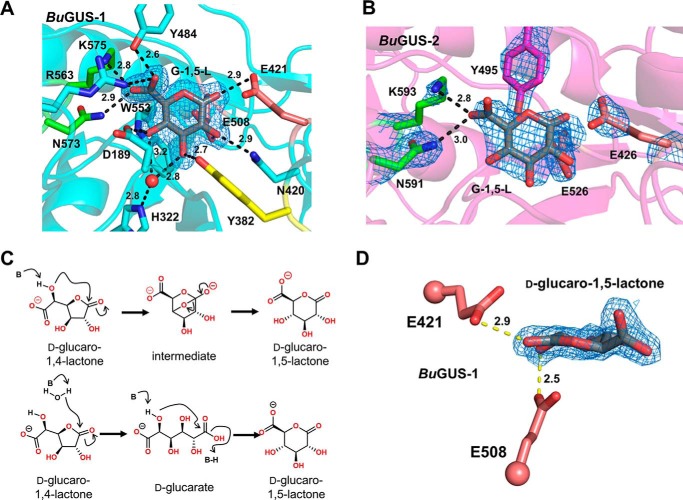
The glycoside hydrolases encoded by the human gut microbiome play an integral role in processing a variety of exogenous and endogenous glycoconjugates. Here we present three structurally and functionally distinct β-glucuronidase (GUS) glycoside hydrolases from a single human gut commensal microbe, Bacteroides uniformis We show using nine crystal structures, biochemical, and biophysical data that whereas these three proteins share similar overall folds, they exhibit different structural features that create three structurally and functionally unique enzyme active sites. Notably, quaternary structure plays an important role in creating distinct active site features that are hard to predict via structural modeling methods. The enzymes display differential processing capabilities toward glucuronic acid-containing polysaccharides and SN-38-glucuronide, a metabolite of the cancer drug irinotecan. We also demonstrate that GUS-specific and nonselective inhibitors exhibit varying potencies toward each enzyme. Together, these data highlight the diversity of GUS enzymes within a single Bacteroides gut commensal and advance our understanding of how structural details impact the specific roles microbial enzymes play in processing drug-glucuronide and glycan substrates.
Keywords: GI tract; bacteroides; carbohydrate metabolism; crystallography; drug reactivation; enzyme structure; glucuronidase; glycoside hydrolase; microbiome; structure-function.
© 2018 Pellock et al.
Conflict of interest statement
M. R. R. is the scientific founder of Symberix, Inc., which is developing microbiome-targeted therapeutics
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
