

Rab34 small GTPase is required for Hedgehog signaling and an early step of ciliary vesicle formation in mouse
- PMID: 30301781
- PMCID: PMC6240299
- DOI: 10.1242/jcs.213710
Rab34 small GTPase is required for Hedgehog signaling and an early step of ciliary vesicle formation in mouse
Abstract
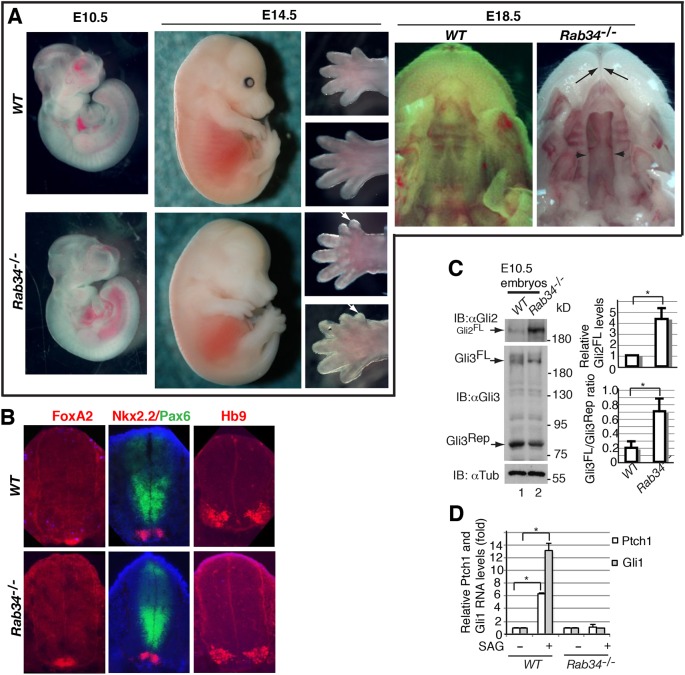
The primary cilium is a microtubule-based organelle that protrudes from the cell surface and plays essential roles in embryonic development. Ciliogenesis begins with the successive fusion of preciliary vesicles to form ciliary vesicles, which then dock onto the distal end of the mother centriole. Rab proteins have been linked to cilia formation in cultured cells, but not yet in vivo In the present study, we demonstrate that endocytic recycling protein Rab34 localizes to cilia, and that its mutation results in significant decrease of ciliogenesis in both cultured cells and mice. Rab34 is required for the successive fusion of preciliary vesicles to generate ciliary vesicles and for the migration of the mother centriole from perinuclear region to plasma membrane. We also show that Rab34 mutant mice exhibit polydactyly, and cleft-lip and -palate. These phenotypes are consistent with observations that nonciliated Rab34 mutant cells fail to respond to Hedgehog signaling and that processing of full-length Gli3 to its C-terminally truncated form is reduced in Rab34 mutant embryos. Therefore, Rab34 is required for an early step of ciliary vesicle formation and Hh signaling in vivo This article has an associated First Person interview with the first author of the paper.
Keywords: Cilia; Ciliary vesicle; Hedgehog; Rab34.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
