

Modernized Tools for Streamlined Genetic Manipulation and Comparative Study of Wild and Diverse Proteobacterial Lineages
- PMID: 30301859
- PMCID: PMC6178617
- DOI: 10.1128/mBio.01877-18
Modernized Tools for Streamlined Genetic Manipulation and Comparative Study of Wild and Diverse Proteobacterial Lineages
Abstract
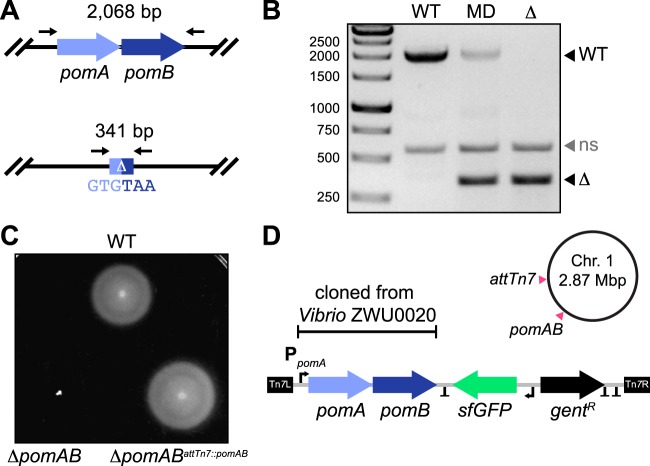
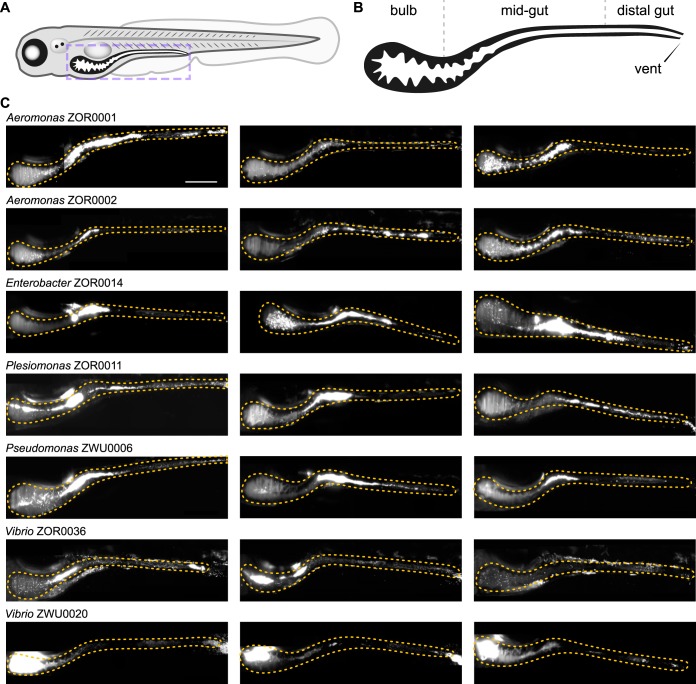
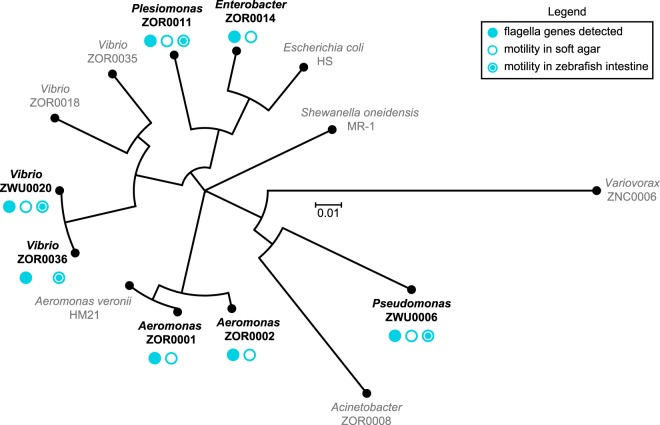
Correlating the presence of bacteria and the genes they carry with aspects of plant and animal biology is rapidly outpacing the functional characterization of naturally occurring symbioses. A major barrier to mechanistic studies is the lack of tools for the efficient genetic manipulation of wild and diverse bacterial isolates. To address the need for improved molecular tools, we used a collection of proteobacterial isolates native to the zebrafish intestinal microbiota as a testbed to construct a series of modernized vectors that expedite genetic knock-in and knockout procedures across lineages. The innovations that we introduce enhance the flexibility of conventional genetic techniques, making it easier to manipulate many different bacterial isolates with a single set of tools. We developed alternative strategies for domestication-free conjugation, designed plasmids with customizable features, and streamlined allelic exchange using visual markers of homologous recombination. We demonstrate the potential of these tools through a comparative study of bacterial behavior within the zebrafish intestine. Live imaging of fluorescently tagged isolates revealed a spectrum of distinct population structures that differ in their biogeography and dominant growth mode (i.e., planktonic versus aggregated). Most striking, we observed divergent genotype-phenotype relationships: several isolates that are predicted by genomic analysis and in vitro assays to be capable of flagellar motility do not display this trait within living hosts. Together, the tools generated in this work provide a new resource for the functional characterization of wild and diverse bacterial lineages that will help speed the research pipeline from sequencing-based correlations to mechanistic underpinnings.IMPORTANCE A great challenge in microbiota research is the immense diversity of symbiotic bacteria with the capacity to impact the lives of plants and animals. Moving beyond correlative DNA sequencing-based studies to define the cellular and molecular mechanisms by which symbiotic bacteria influence the biology of their hosts is stalling because genetic manipulation of new and uncharacterized bacterial isolates remains slow and difficult with current genetic tools. Moreover, developing tools de novo is an arduous and time-consuming task and thus represents a significant barrier to progress. To address this problem, we developed a suite of engineering vectors that streamline conventional genetic techniques by improving postconjugation counterselection, modularity, and allelic exchange. Our modernized tools and step-by-step protocols will empower researchers to investigate the inner workings of both established and newly emerging models of bacterial symbiosis.
Keywords: Tn7; allelic exchange; bacterial genetics; conjugation; counterselection; domestication; genetic manipulation; microbiome; modular tools; proteobacteria; symbiosis; zebrafish.
Copyright © 2018 Wiles et al.
Figures

References
-
- McFall-Ngai M, Hadfield MG, Bosch TCG, Carey HV, Domazet-Lošo T, Douglas AE, Dubilier N, Eberl G, Fukami T, Gilbert SF, Hentschel U, King N, Kjelleberg S, Knoll AH, Kremer N, Mazmanian SK, Metcalf JL, Nealson K, Pierce NE, Rawls JF, Reid A, Ruby EG, Rumpho M, Sanders JG, Tautz D, Wernegreen JJ. 2013. Animals in a bacterial world, a new imperative for the life sciences. Proc Natl Acad Sci U S A 110:3229–3236. doi:10.1073/pnas.1218525110. - DOI - PMC - PubMed
-
- Cox LM, Yamanishi S, Sohn J, Alekseyenko AV, Leung JM, Cho I, Kim SG, Li H, Gao Z, Mahana D, Zárate Rodriguez JG, Rogers AB, Robine N, Loke P, Blaser MJ. 2014. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell 158:705–721. doi:10.1016/j.cell.2014.05.052. - DOI - PMC - PubMed
-
- Kostic AD, Gevers D, Siljander H, Vatanen T, Hyötyläinen T, Hämäläinen A-M, Peet A, Tillmann V, Pöhö P, Mattila I, Lähdesmäki H, Franzosa EA, Vaarala O, de Goffau M, Harmsen H, Ilonen J, Virtanen SM, Clish CB, Orešič M, Huttenhower C, Knip M, Diabimmune Study Group, Xavier RJ. 2015. The dynamics of the human infant gut microbiome in development and in progression toward type 1 diabetes. Cell Host Microbe 17:260–273. doi:10.1016/j.chom.2015.01.001. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
