

Cryo-EM structure of the hibernating Thermus thermophilus 100S ribosome reveals a protein-mediated dimerization mechanism
- PMID: 30301898
- PMCID: PMC6177447
- DOI: 10.1038/s41467-018-06724-x
Cryo-EM structure of the hibernating Thermus thermophilus 100S ribosome reveals a protein-mediated dimerization mechanism
Abstract
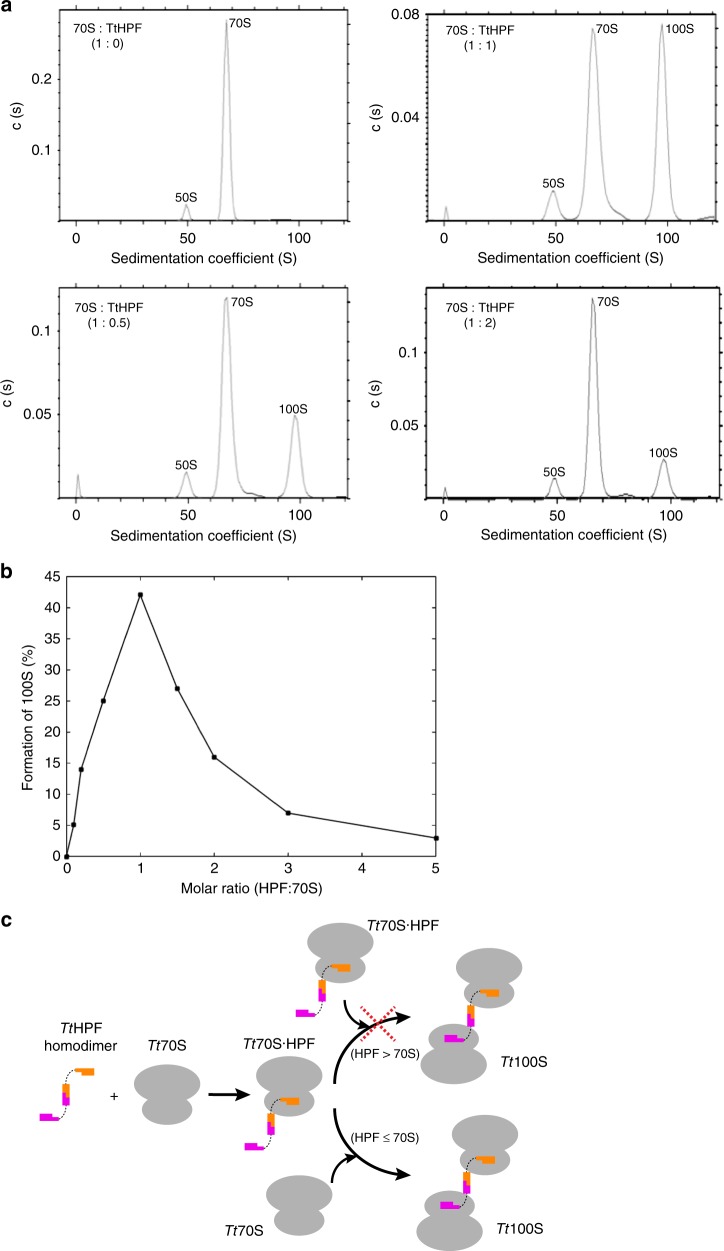
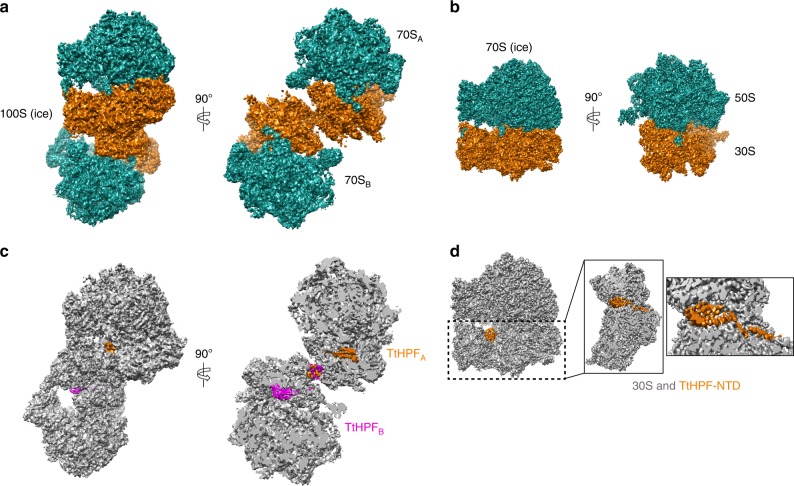
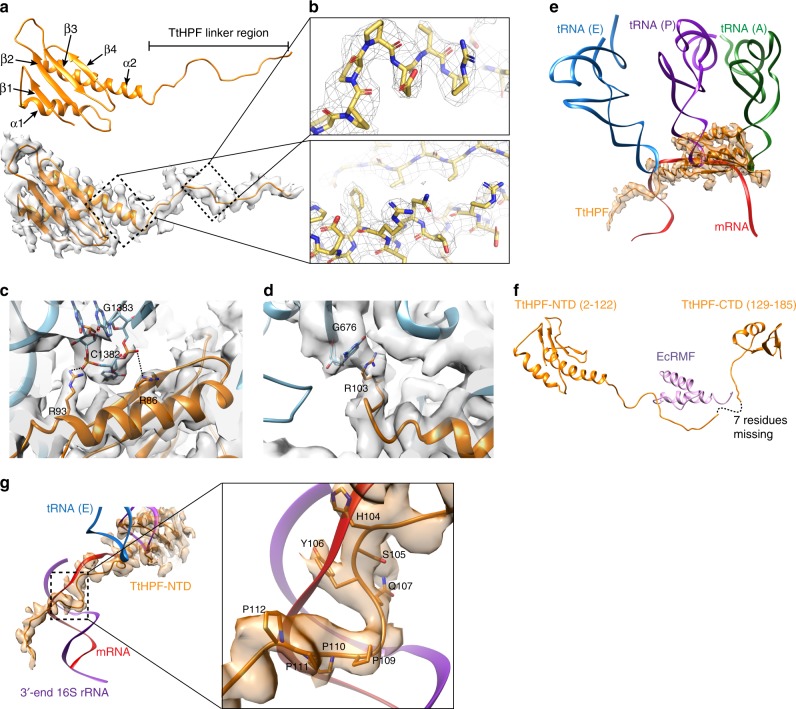
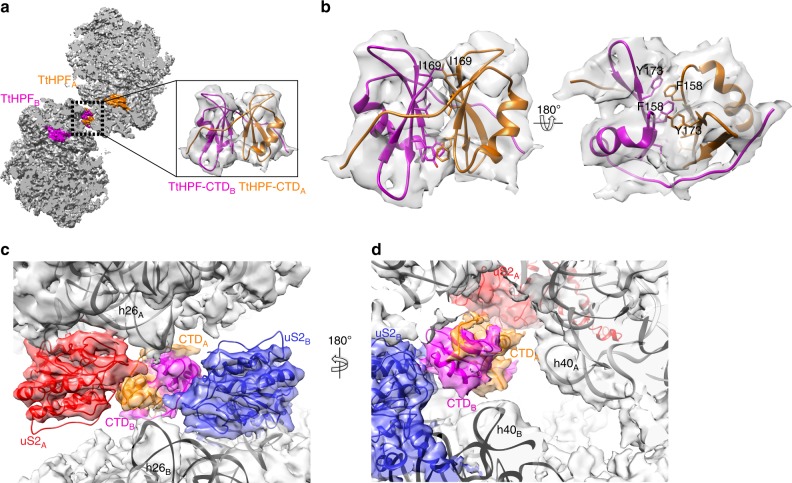
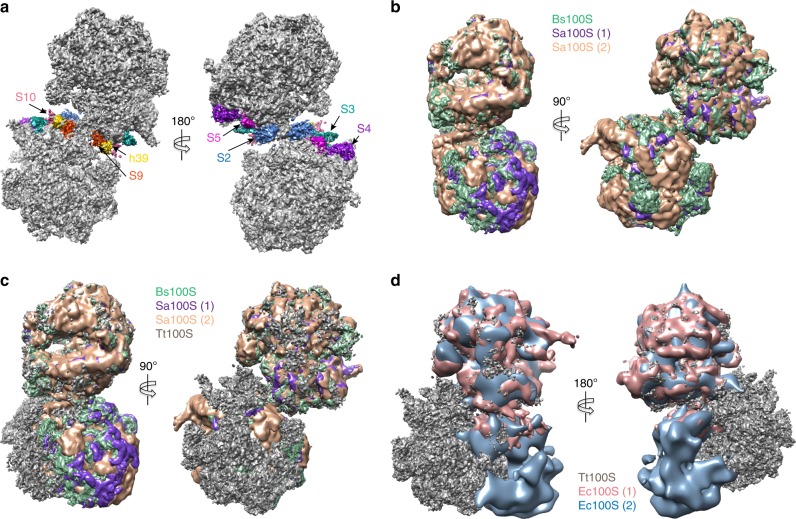
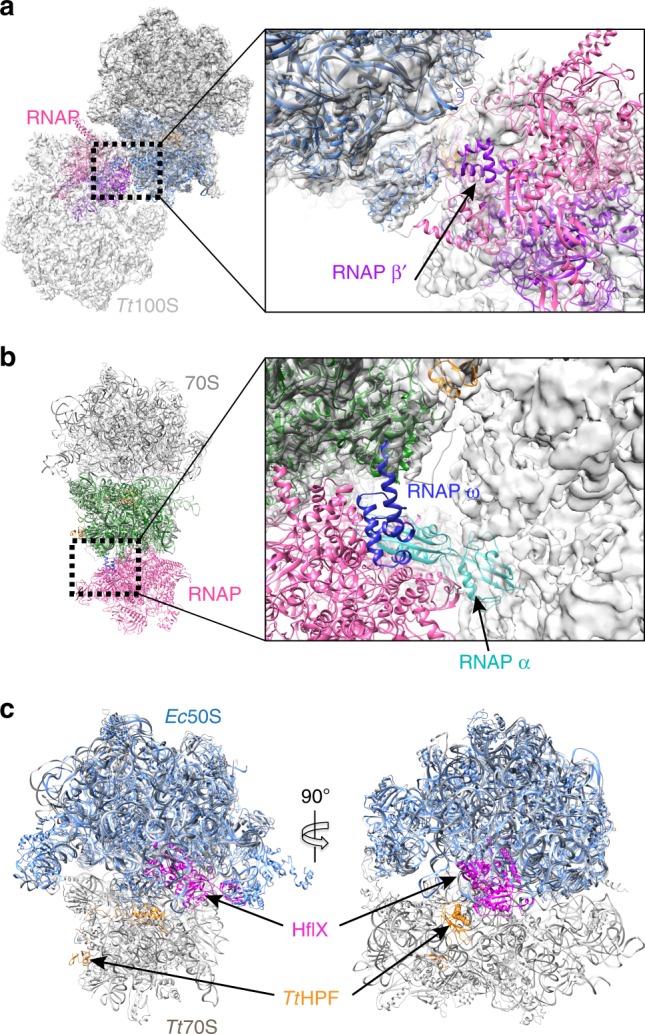
In response to cellular stresses bacteria conserve energy by dimerization of ribosomes into inactive hibernating 100S ribosome particles. Ribosome dimerization in Thermus thermophilus is facilitated by hibernation-promoting factor (TtHPF). In this study we demonstrate high sensitivity of Tt100S formation to the levels of TtHPF and show that a 1:1 ratio leads to optimal dimerization. We report structures of the T. thermophilus 100S ribosome determined by cryo-electron microscopy to average resolutions of 4.13 Å and 4.57 Å. In addition, we present a 3.28 Å high-resolution cryo-EM reconstruction of a 70S ribosome from a hibernating ribosome dimer and reveal a role for the linker region connecting the TtHPF N- and C-terminal domains in translation inhibition by preventing Shine-Dalgarno duplex formation. Our work demonstrates that species-specific differences in the dimerization interface govern the overall conformation of the 100S ribosome particle and that for Thermus thermophilus no ribosome-ribosome interactions are involved in the interface.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Stress response as implemented by hibernating ribosomes: a structural overview.FEBS J. 2019 Sep;286(18):3558-3565. doi: 10.1111/febs.14968. Epub 2019 Jul 15. FEBS J. 2019. PMID: 31230411 Free PMC article. Review.
-
Structure of the Bacillus subtilis hibernating 100S ribosome reveals the basis for 70S dimerization.EMBO J. 2017 Jul 14;36(14):2061-2072. doi: 10.15252/embj.201696189. Epub 2017 May 3. EMBO J. 2017. PMID: 28468753 Free PMC article.
-
Structure of a hibernating 100S ribosome reveals an inactive conformation of the ribosomal protein S1.Nat Microbiol. 2018 Oct;3(10):1115-1121. doi: 10.1038/s41564-018-0237-0. Epub 2018 Sep 3. Nat Microbiol. 2018. PMID: 30177741
-
A general mechanism of ribosome dimerization revealed by single-particle cryo-electron microscopy.Nat Commun. 2017 Sep 28;8(1):722. doi: 10.1038/s41467-017-00718-x. Nat Commun. 2017. PMID: 28959045 Free PMC article.
-
The ribosome through the looking glass.Angew Chem Int Ed Engl. 2003 Aug 4;42(30):3464-86. doi: 10.1002/anie.200200544. Angew Chem Int Ed Engl. 2003. PMID: 12900959 Review.
Cited by
-
Stress response as implemented by hibernating ribosomes: a structural overview.FEBS J. 2019 Sep;286(18):3558-3565. doi: 10.1111/febs.14968. Epub 2019 Jul 15. FEBS J. 2019. PMID: 31230411 Free PMC article. Review.
-
Interpretation of anomalously long crosslinks in ribosome crosslinking reveals the ribosome interaction in stationary phase E. coli.RSC Chem Biol. 2022 May 16;3(7):886-894. doi: 10.1039/d2cb00101b. eCollection 2022 Jul 6. RSC Chem Biol. 2022. PMID: 35866168 Free PMC article.
-
Draft genome sequences of Thermus thermophilus strains isolated from Japanese hot springs.Microbiol Resour Announc. 2025 Jul 10;14(7):e0055925. doi: 10.1128/mra.00559-25. Epub 2025 Jun 20. Microbiol Resour Announc. 2025. PMID: 40646720 Free PMC article.
-
Purification of Hibernating and Active C- Ribosomes from Zinc-Starved Mycobacteria.Methods Mol Biol. 2021;2314:151-166. doi: 10.1007/978-1-0716-1460-0_5. Methods Mol Biol. 2021. PMID: 34235651 Free PMC article.
-
Ribosome hibernation: a new molecular framework for targeting nonreplicating persisters of mycobacteria.Microbiology (Reading). 2021 Feb;167(2):001035. doi: 10.1099/mic.0.001035. Microbiology (Reading). 2021. PMID: 33555244 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
