

The neural mechanisms and circuitry of the pair bond
- PMID: 30301953
- PMCID: PMC6283620
- DOI: 10.1038/s41583-018-0072-6
The neural mechanisms and circuitry of the pair bond
Abstract
Love is one of our most powerful emotions, inspiring some of the greatest art, literature and conquests of human history. Although aspects of love are surely unique to our species, human romantic relationships are displays of a mating system characterized by pair bonding, likely built on ancient foundational neural mechanisms governing individual recognition, social reward, territorial behaviour and maternal nurturing. Studies in monogamous prairie voles and mice have revealed precise neural mechanisms regulating processes essential for the pair bond. Here, we discuss current viewpoints on the biology underlying pair bond formation, its maintenance and associated behaviours from neural and evolutionary perspectives.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
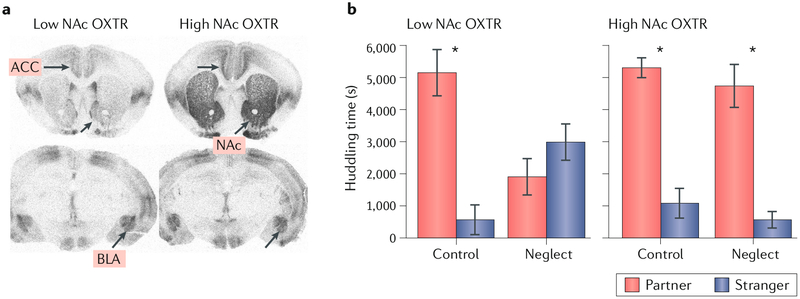
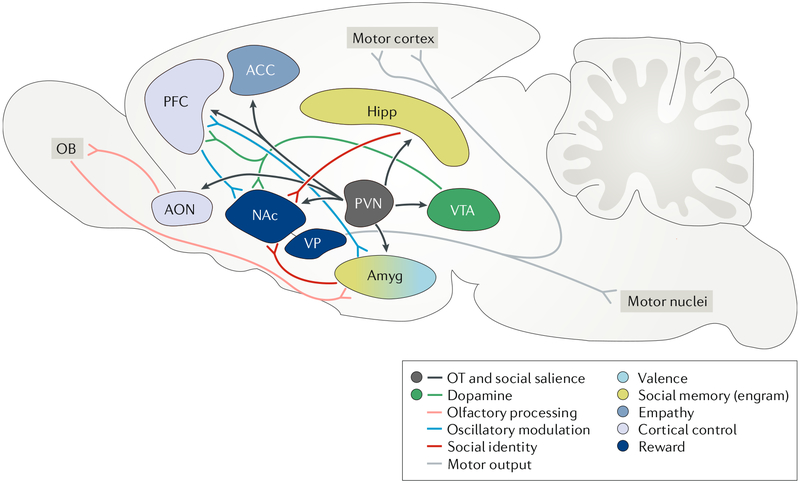
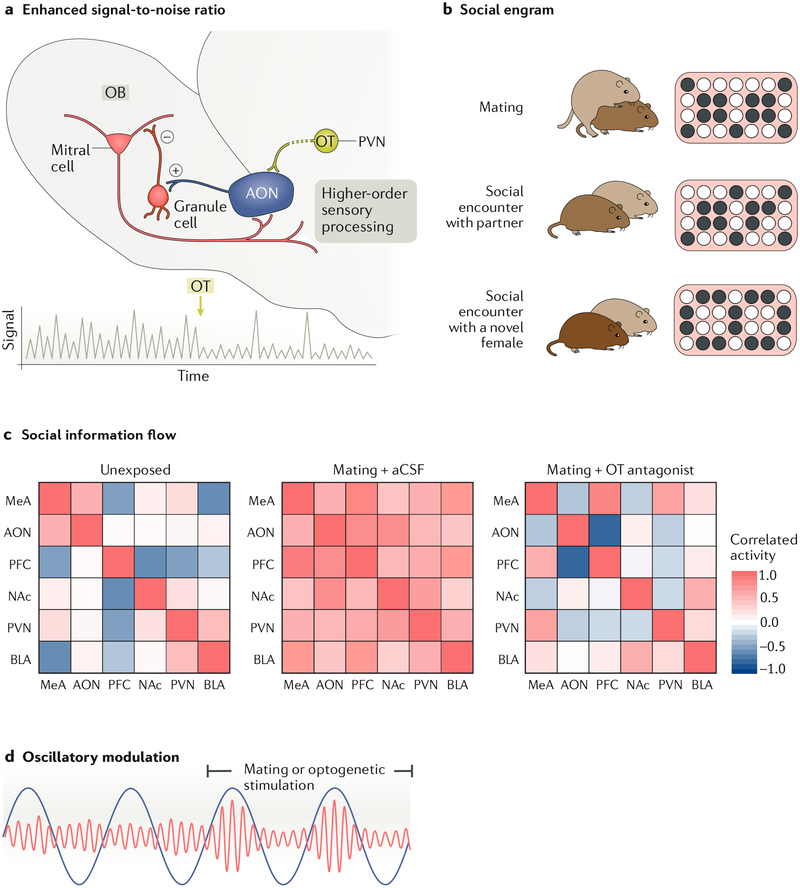
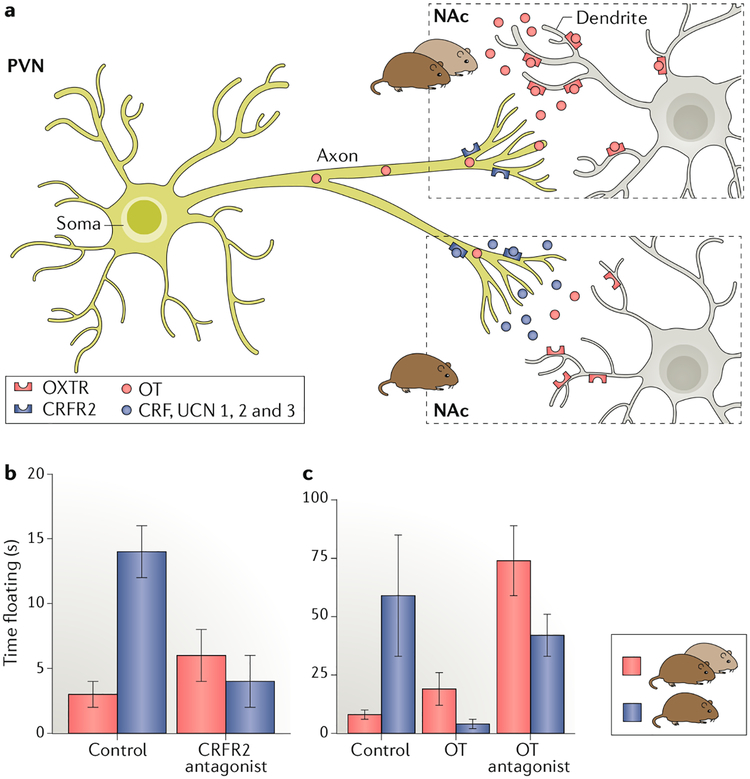
References
-
- Young LJ Being human: love: neuroscience reveals all. Nature 457, 148 (2009). - PubMed
-
- Bull CM Monogamy in lizards. Behav. Processes 51, 7–20 (2000). - PubMed
-
- Whiteman EA & Cote IM Monogamy in marine fishes. Biol. Rev. Camb. Philos. Soc 79, 351–375 (2004). - PubMed
-
- Dunn PO, Whittingham LA & Pitcher TE Mating systems, sperm competition, and the evolution of sexual dimorphism in birds. Evolution 55, 161–175 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
