

Basal and starvation-induced autophagy mediates parasite survival during intraerythrocytic stages of Plasmodium falciparum
- PMID: 30302277
- PMCID: PMC6170468
- DOI: 10.1038/s41420-018-0107-9
Basal and starvation-induced autophagy mediates parasite survival during intraerythrocytic stages of Plasmodium falciparum
Erratum in
-
Erratum: Publisher Correction: articles initially published in wrong volume.Cell Death Discov. 2019 Jul 10;5:116. doi: 10.1038/s41420-019-0186-2. eCollection 2019. Cell Death Discov. 2019. PMID: 31312525 Free PMC article.
Abstract
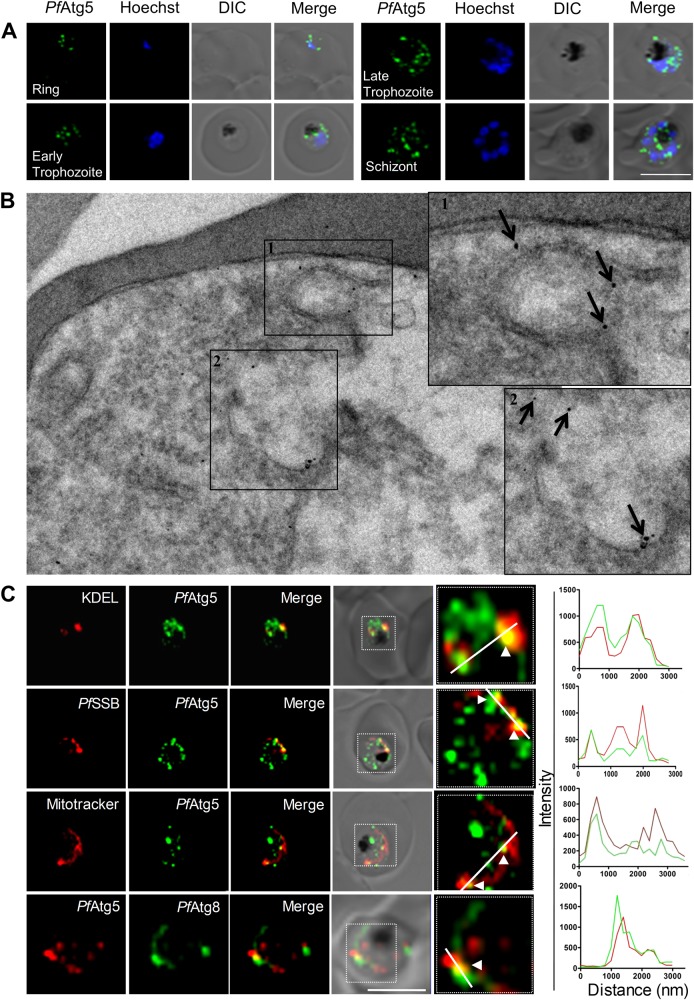
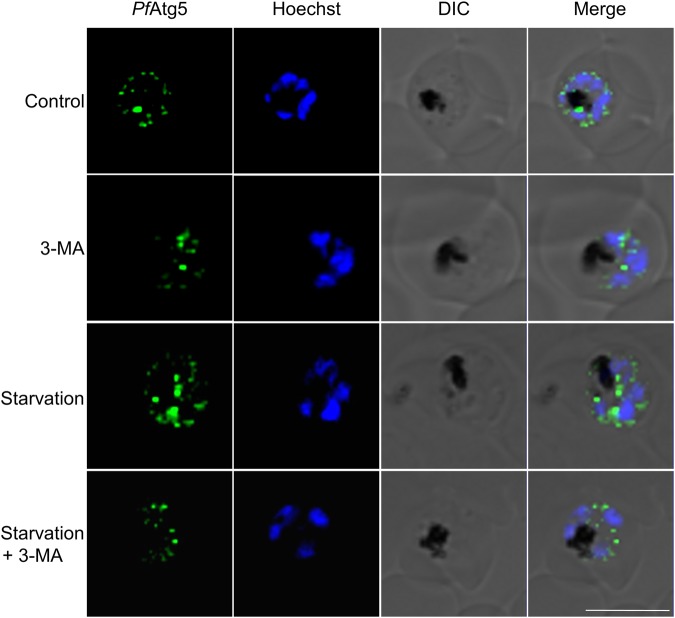
The precise role of autophagy in P. falciparum remains largely unknown. Although a limited number of autophagy genes have been identified in this apicomplexan, only PfAtg8 has been characterized to a certain extent. On the basis of the expression levels of PfAtg8 and the putative PfAtg5, we report that the basal autophagy in this parasite is quite robust and mediates not only the intraerythrocytic development but also fresh invasion of red blood cells (RBCs) in the subsequent cycles. We demonstrate that the basal autophagy responds to both inducers and inhibitors of autophagy. In addition, the parasite survival upon starvation is temporally governed by the autophagy status. Brief periods of starvation, which induces autophagy, help survival while prolonged starvation decreases autophagy leading to stalled parasite growth and reduced invasion. Thus, starvation-induced autophagy is context dependent. Importantly, we report characterization of another autophagy marker in this parasite, the putative PfAtg5 (Pf3D7_1430400). PfAtg5 is expressed in all the intraerythrocytic stages and partially colocalizes with ER, mitochondria, apicoplast and PfAtg8. It is also present on the double membrane bound vesicles. Altogether, these studies pave way for the detailed dissection of P. falciparum autophagy machinery and insights into molecular and functional characterization of its players for developing new therapeutics as antimalarials.
Conflict of interest statement
The authors declare no potential conflict of interest.
Figures
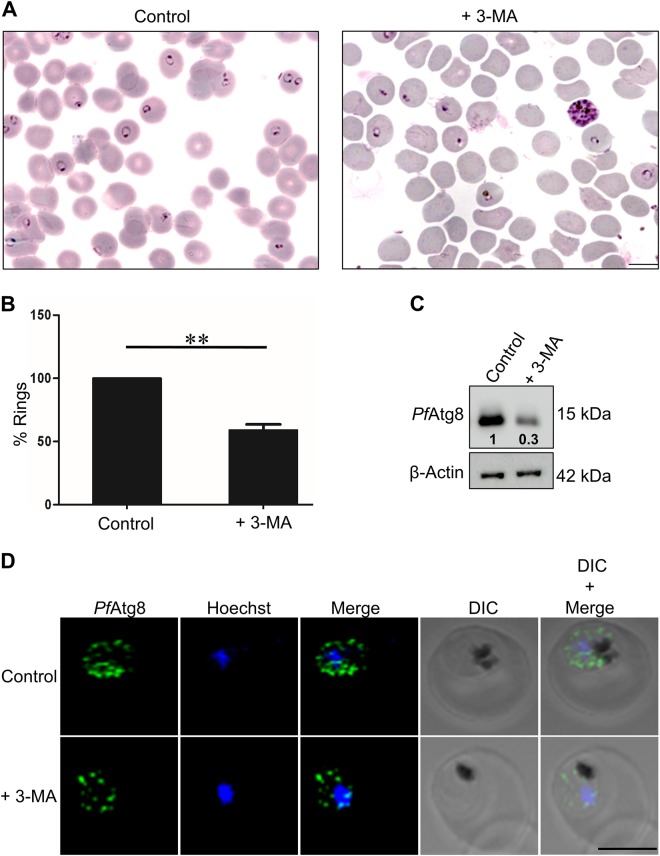
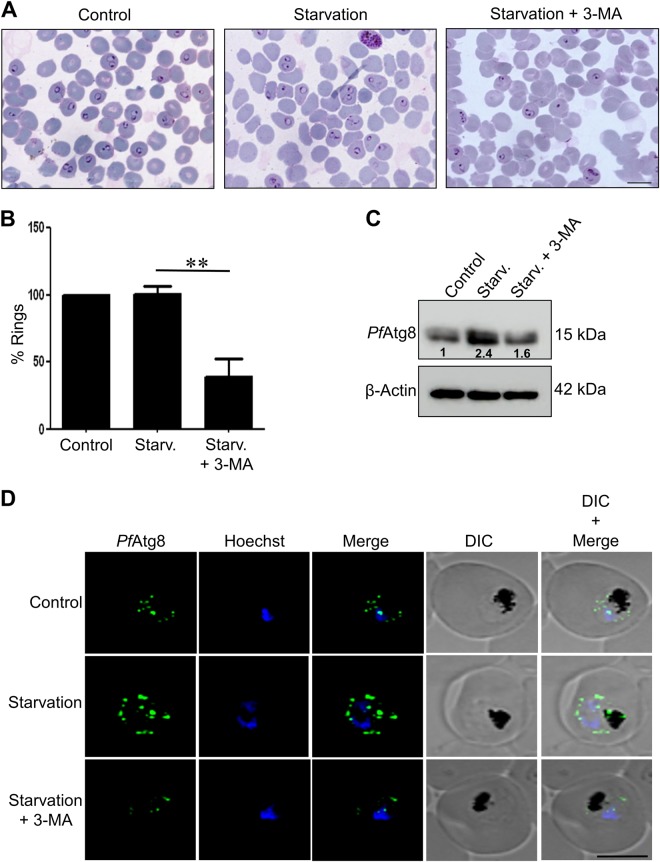
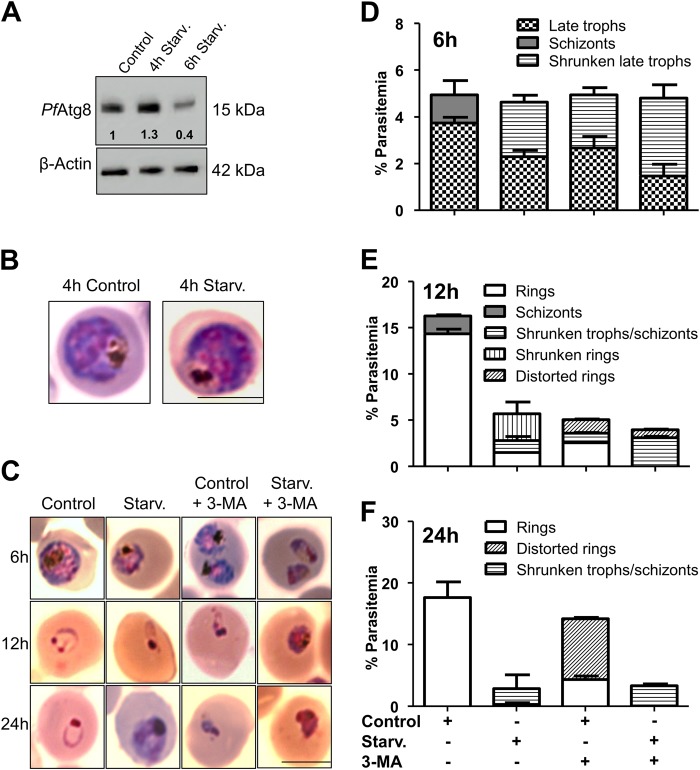
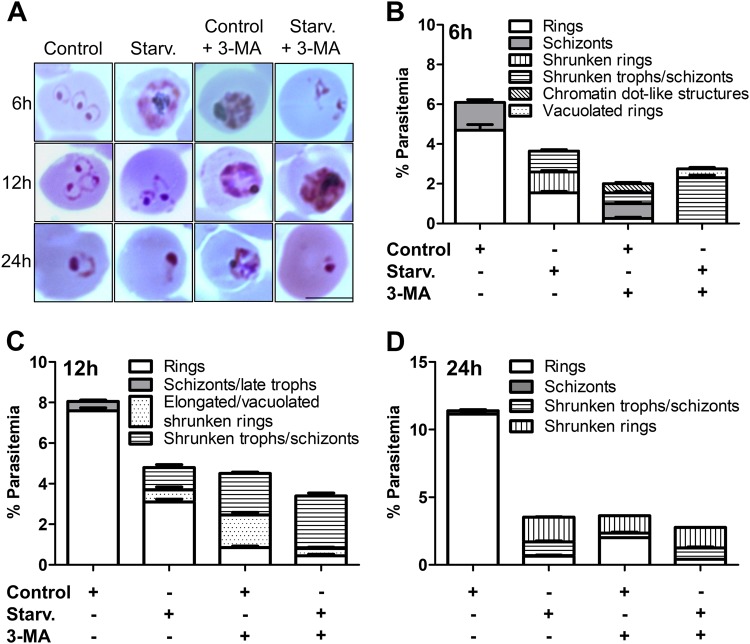
References
Grants and funding
LinkOut - more resources
Full Text Sources
