

Review
doi: 10.1152/physiol.00029.2018.
The Difference δ-Cells Make in Glucose Control
Affiliations
- PMID: 30303773
- PMCID: PMC6347098
- DOI: 10.1152/physiol.00029.2018
Item in Clipboard
Review
The Difference δ-Cells Make in Glucose Control
Physiology (Bethesda).
.
Abstract
The role of beta and α-cells to glucose control are established, but the physiological role of δ-cells is poorly understood. Delta-cells are ideally positioned within pancreatic islets to modulate insulin and glucagon secretion at their source. We review the evidence for a negative feedback loop between delta and β-cells that determines the blood glucose set point and suggest that local δ-cell-mediated feedback stabilizes glycemic control.
Figures
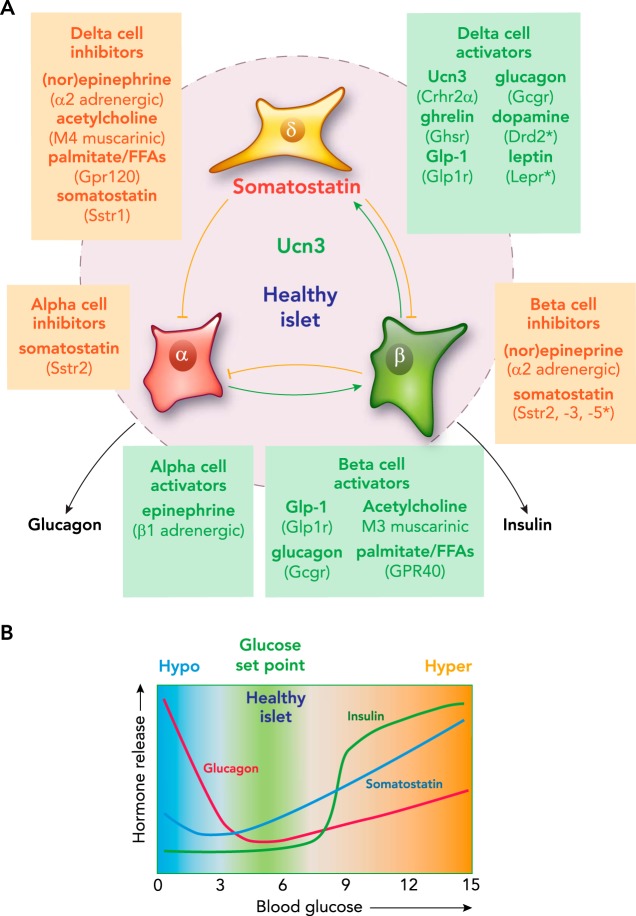
Pancreatic δ-cells A: pancreatic δ-cells receive input from numerous paracrine, endocrine, and neural inputs, and translate this into appropriate inhibition of glucagon and insulin release by α- and β-cells. Select stimulatory and inhibitory inputs are given for each of the islet cell types. B: schematic representation of the profiles of insulin, glucagon, and somatostatin secretion as a function of blood glucose.
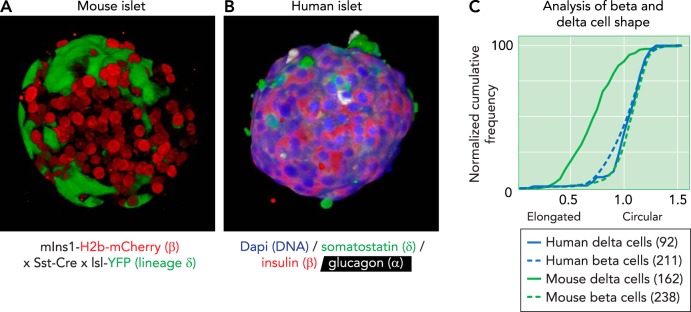
Projection of a 3D reconstruction of a pancreatic islet from a transgenic reporter strain A: projection of a 3D reconstruction of a pancreatic islet from a transgenic reporter strain, captured by confocal microscope. β-Cells are visualized by the nuclear expression of an mCherry under control of the Ins1 promoter. δ-Cells are visualized by the expression of Cre recombinase under control of the somatostatin (Sst) promotor, which leads to their irreversible expression of yellow fluorescent protein (YFP; green). B: projection of a 3D reconstruction of a human pancreatic islet, captured by confocal microscope and stained for somatostatin (green), insulin (red), and glucagon (white). Nuclei (dapi) are counterstained in blue. Note how the human δ-cells are notably more compact compared with the axon-like mouse δ-cells. C: the difference in morphology of mouse and human δ-cells and β-cells from the same islet quantified their circularity, defined as the normalized ratio of the area over the perimeter of the cell outline. Each cell outline was determined in Nikon Elements, and circularity was calculated as . A value of 1 indicates a perfect circle. Mouse δ-cells stand out for their elongated morphology, which manifests as a significant reduction in circularity, compared with β-cells, whereas human δ-cells are similarly compact to human β-cells. Numbers in-between parentheses reflect the number of cells quantified from 3D confocal reconstructions of intact islets from two individual subjects for each species.
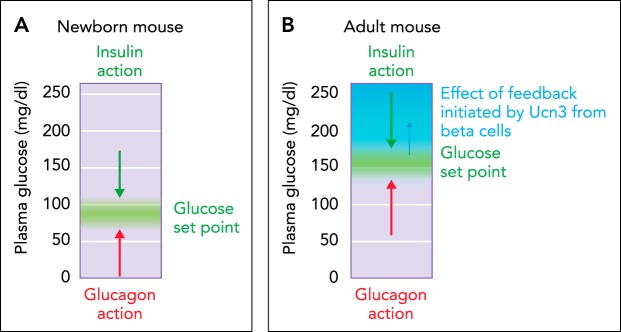
The homeostatic set point for glucose A: in the absence of UCN3 in young neonatal mice, the homeostatic set point for glucose is determined by the balance between insulin and glucagon action. B: after the onset of UCN3 expression in mouse β-cells, β-cell activation leads to the co-secretion of UCN3 with insulin. This activates feedback inhibition that curbs insulin secretion and effectively reduces insulin action.
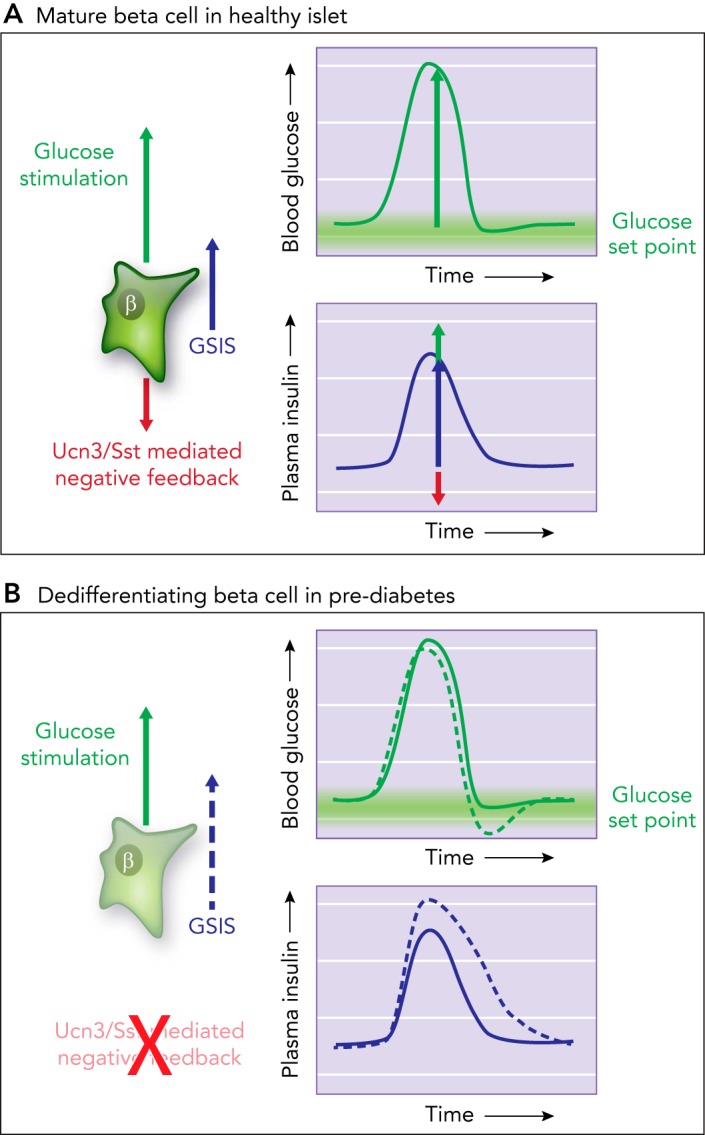
Model of tonic feedback inhibition on β- and α-cells A: model how tonic feedback inhibition on β- and α-cells ensures the timely attenuation of insulin (or glucagon) secretion as glucose is restored to its homeostatic equilibrium. B: when this local feedback breaks down, insulin secretion is des-inhibited. This prolongs insulin action, which causes glucose values to overshoot their glucose set point and contributes to hyperglycemia in diabetes.
References
-
- Adriaenssens AE, Svendsen B, Lam BY, Yeo GS, Holst JJ, Reimann F, Gribble FM. Transcriptomic profiling of pancreatic alpha, beta and δ-cell populations identifies δ-cells as a principal target for ghrelin in mouse islets. Diabetologia 59: 2156–2165, 2016. doi:10.1007/s00125-016-4033-1. - DOI - PMC - PubMed
-
- Benner C, van der Meulen T, Cacéres E, Tigyi K, Donaldson CJ, Huising MO. The transcriptional landscape of mouse β-cells compared to human β-cells reveals notable species differences in long non-coding RNA and protein-coding gene expression. BMC Genomics 15: 620, 2014. doi:10.1186/1471-2164-15-620. - DOI - PMC - PubMed