

Most Sinorhizobium meliloti Extracytoplasmic Function Sigma Factors Control Accessory Functions
- PMID: 30305320
- PMCID: PMC6180224
- DOI: 10.1128/mSphereDirect.00454-18
Most Sinorhizobium meliloti Extracytoplasmic Function Sigma Factors Control Accessory Functions
Abstract
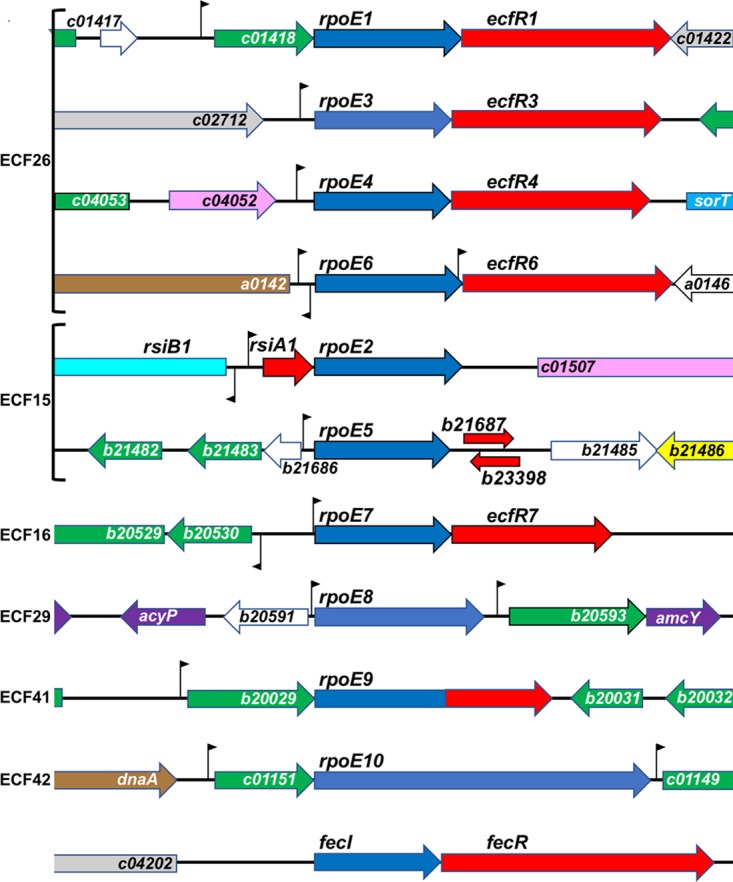
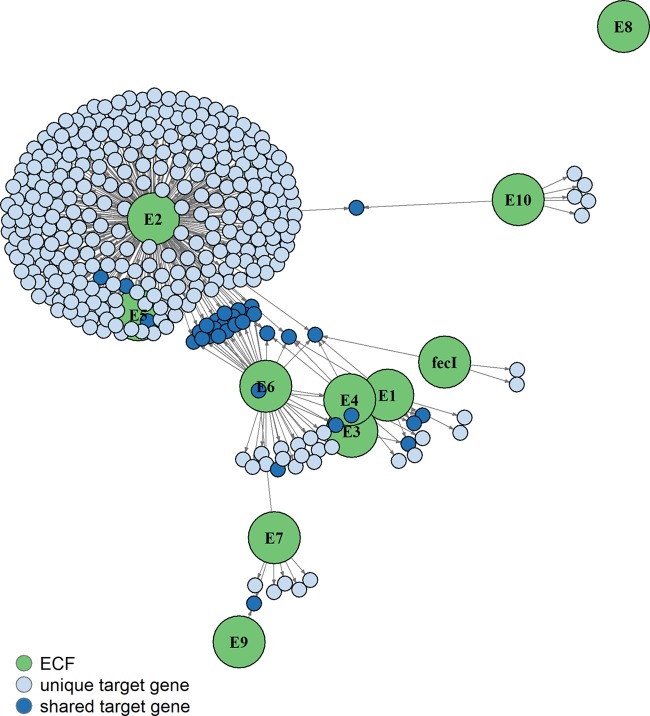
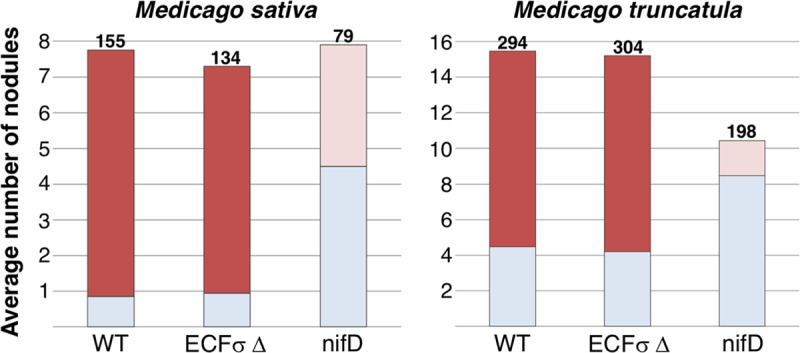
Bacteria must sense alterations in their environment and respond with changes in function and/or structure in order to cope. Extracytoplasmic function sigma factors (ECF σs) modulate transcription in response to cellular and environmental signals. The symbiotic nitrogen-fixing alphaproteobacterium Sinorhizobium meliloti carries genes for 11 ECF-like σs (RpoE1 to -E10 and FecI). We hypothesized that some of these play a role in mediating the interaction between the bacterium and its plant symbiotic partner. The bacterium senses changes in its immediate environment as it establishes contact with the plant root, initiates invasion of the plant as the root nodule is formed, traverses several root cell layers, and enters plant cortical cells via endocytosis. We used genetics, transcriptomics, and functionality to characterize the entire S. meliloti cohort of ECF σs. We discovered new targets for individual σs, confirmed others by overexpressing individual ECF σs, and identified or confirmed putative promoter motifs for nine of them. We constructed precise deletions of each ECF σ gene and its demonstrated or putative anti-σ gene and also a strain in which all 11 ECF σ and anti-σ genes were deleted. This all-ECF σ deletion strain showed no major defects in free-living growth, in Biolog Phenotype MicroArray assays, or in response to multiple stresses. None of the ECF σs were required for symbiosis on the host plants Medicago sativa and Medicago truncatula: the strain deleted for all ECF σ and anti-σ genes was symbiotically normal.IMPORTANCE Fixed (reduced) soil nitrogen plays a critical role in soil fertility and successful food growth. Much soil fertility relies on symbiotic nitrogen fixation: the bacterial partner infects the host plant roots and reduces atmospheric dinitrogen in exchange for host metabolic fuel, a process that involves complex interactions between the partners mediated by changes in gene expression in each partner. Here we test the roles of a family of 11 extracytoplasmic function (ECF) gene regulatory proteins (sigma factors [σs]) that interact with RNA polymerase to determine if they play a significant role in establishing a nitrogen-fixing symbiosis or in responding to various stresses, including cell envelope stress. We discovered that symbiotic nitrogen fixation occurs even when all 11 of these regulatory genes are deleted, that most ECF sigma factors control accessory functions, and that none of the ECF sigma factors are required to survive envelope stress.
Keywords: Rhizobium; Sinorhizobium; microarrays; sigma factors; symbiosis.
Copyright © 2018 Lang et al.
Figures


Similar articles
-
Contributions of Sinorhizobium meliloti Transcriptional Regulator DksA to Bacterial Growth and Efficient Symbiosis with Medicago sativa.J Bacteriol. 2016 Apr 14;198(9):1374-83. doi: 10.1128/JB.00013-16. Print 2016 May. J Bacteriol. 2016. PMID: 26883825 Free PMC article.
-
Novel Genes and Regulators That Influence Production of Cell Surface Exopolysaccharides in Sinorhizobium meliloti.J Bacteriol. 2018 Jan 10;200(3):e00501-17. doi: 10.1128/JB.00501-17. Print 2018 Feb 1. J Bacteriol. 2018. PMID: 29158240 Free PMC article.
-
Sinorhizobium medicae WSM419 Genes That Improve Symbiosis between Sinorhizobium meliloti Rm1021 and Medicago truncatula Jemalong A17 and in Other Symbiosis Systems.Appl Environ Microbiol. 2021 Jul 13;87(15):e0300420. doi: 10.1128/AEM.03004-20. Epub 2021 Jul 13. Appl Environ Microbiol. 2021. PMID: 33990306 Free PMC article.
-
How rhizobial symbionts invade plants: the Sinorhizobium-Medicago model.Nat Rev Microbiol. 2007 Aug;5(8):619-33. doi: 10.1038/nrmicro1705. Nat Rev Microbiol. 2007. PMID: 17632573 Free PMC article. Review.
-
Rhizobium meliloti exopolysaccharides: synthesis and symbiotic function.Gene. 1996 Nov 7;179(1):141-6. doi: 10.1016/s0378-1119(96)00322-8. Gene. 1996. PMID: 8955640 Review.
Cited by
-
Rhizopine biosensors for plant-dependent control of bacterial gene expression.Environ Microbiol. 2023 Feb;25(2):383-396. doi: 10.1111/1462-2920.16288. Epub 2022 Dec 4. Environ Microbiol. 2023. PMID: 36428208 Free PMC article.
-
Rhizobium etli CFN42 and Sinorhizobium meliloti 1021 bioinformatic transcriptional regulatory networks from culture and symbiosis.Front Bioinform. 2024 Aug 28;4:1419274. doi: 10.3389/fbinf.2024.1419274. eCollection 2024. Front Bioinform. 2024. PMID: 39263245 Free PMC article.
-
Analysis of Ensifer aridi Mutants Affecting Regulation of Methionine, Trehalose, and Inositol Metabolisms Suggests a Role in Stress Adaptation and Symbiosis Development.Microorganisms. 2022 Jan 26;10(2):298. doi: 10.3390/microorganisms10020298. Microorganisms. 2022. PMID: 35208753 Free PMC article.
-
Ancestral zinc-finger bearing protein MucR in alpha-proteobacteria: A novel xenogeneic silencer?Comput Struct Biotechnol J. 2020 Nov 19;18:3623-3631. doi: 10.1016/j.csbj.2020.11.028. eCollection 2020. Comput Struct Biotechnol J. 2020. PMID: 33304460 Free PMC article. Review.
-
A MoClo-Compatible Toolbox of ECF Sigma Factor-Based Regulatory Switches for Proteobacterial Chassis.Biodes Res. 2024 Feb 21;6:0025. doi: 10.34133/bdr.0025. eCollection 2024. Biodes Res. 2024. PMID: 38384496 Free PMC article.
References
-
- Long SR, Kahn ML, Seefeldt L, Tsay Y-F, Kopriva S. 2015. Nitrogen and sulfur, p 711–768. In Buchanan BB, Gruissem W, Jones RL (ed), Biochemistry and molecular biology of plants. Wiley Blackwell, Oxford, United Kingdom.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
