

Impact of parallel fiber to Purkinje cell long-term depression is unmasked in absence of inhibitory input
- PMID: 30306129
- PMCID: PMC6170036
- DOI: 10.1126/sciadv.aas9426
Impact of parallel fiber to Purkinje cell long-term depression is unmasked in absence of inhibitory input
Abstract
Pavlovian eyeblink conditioning has been used extensively to study the neural mechanisms underlying associative and motor learning. During this simple learning task, memory formation takes place at Purkinje cells in defined areas of the cerebellar cortex, which acquire a strong temporary suppression of their activity during conditioning. Yet, it is unknown which neuronal plasticity mechanisms mediate this suppression. Two potential mechanisms include long-term depression of parallel fiber to Purkinje cell synapses and feed-forward inhibition by molecular layer interneurons. We show, using a triple transgenic approach, that only concurrent disruption of both these suppression mechanisms can severely impair conditioning, highlighting that both processes can compensate for each other's deficits.
Figures
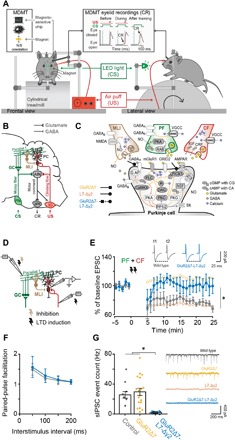
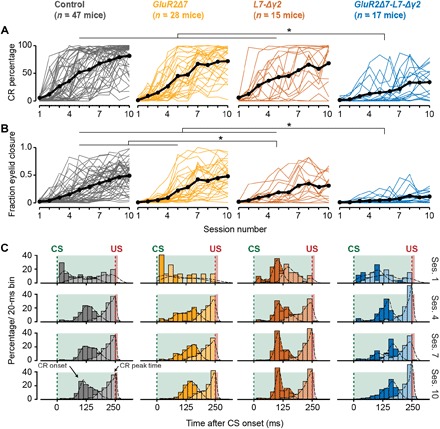
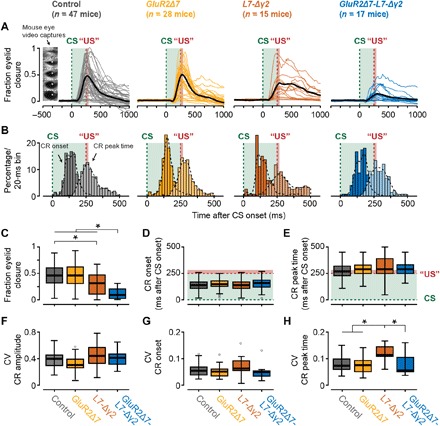
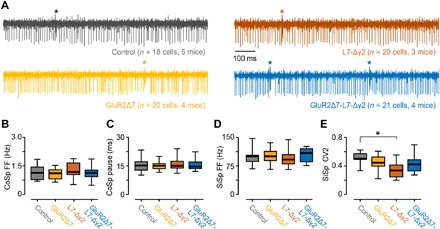
References
-
- H. J. Boele, M. M. Ten Brinke, C. I. De Zeeuw, Classical conditioning of timed motor responses: Neural coding in cerebellar cortex and cerebellar nuclei, in The Neural Codes of the Cerebellum, D. H. Heck, Ed. (Academic Press, 2015), pp. 53–96.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
