

Neurocardiac regulation: from cardiac mechanisms to novel therapeutic approaches
- PMID: 30307615
- PMCID: PMC7496613
- DOI: 10.1113/JP276962
Neurocardiac regulation: from cardiac mechanisms to novel therapeutic approaches
Abstract
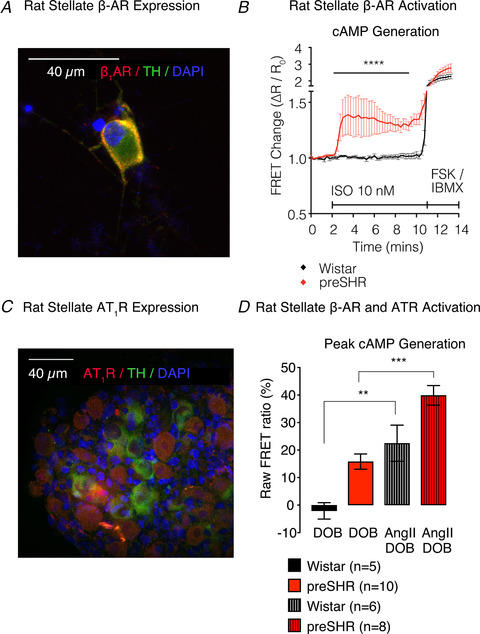
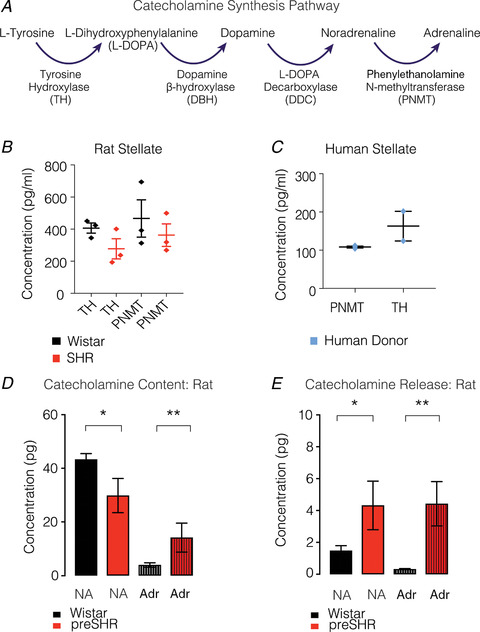
Cardiac sympathetic overactivity is a well-established contributor to the progression of neurogenic hypertension and heart failure, yet the underlying pathophysiology remains unclear. Recent studies have highlighted the importance of acutely regulated cyclic nucleotides and their effectors in the control of intracellular calcium and exocytosis. Emerging evidence now suggests that a significant component of sympathetic overactivity and enhanced transmission may arise from impaired cyclic nucleotide signalling, resulting from compromised phosphodiesterase activity, as well as alterations in receptor-coupled G-protein activation. In this review, we address some of the key cellular and molecular pathways that contribute to sympathetic overactivity in hypertension and discuss their potential for therapeutic targeting.
Keywords: Autonomic Nervous System; Cardiovascular Disease; Cyclic Nucleotide; Hypertension; Intracellular Calcium; Protein Kinase; Sympathetic Ganglion; Sympathetic Nervous System.
© 2018 The Authors. The Journal of Physiology published by John Wiley & Sons Ltd on behalf of The Physiological Society.
Figures
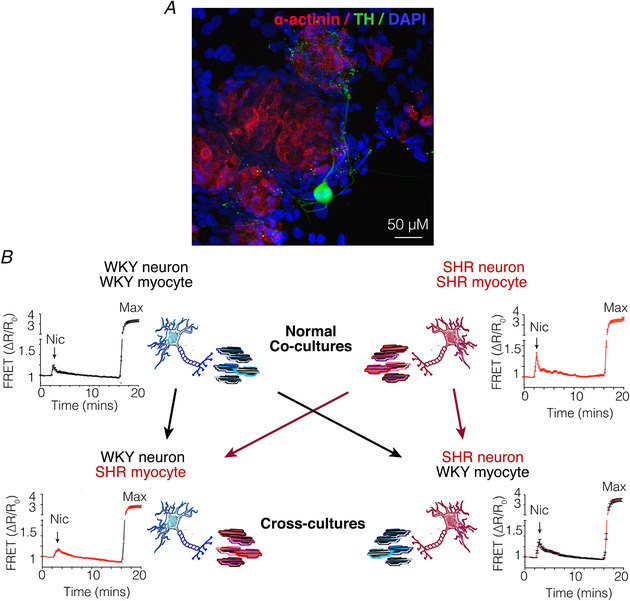
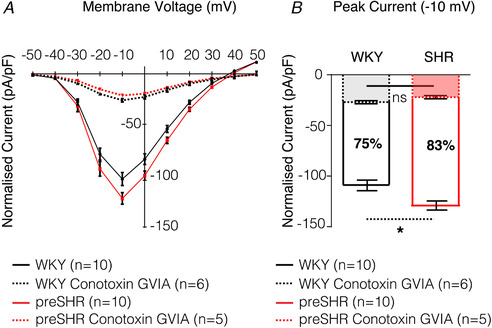
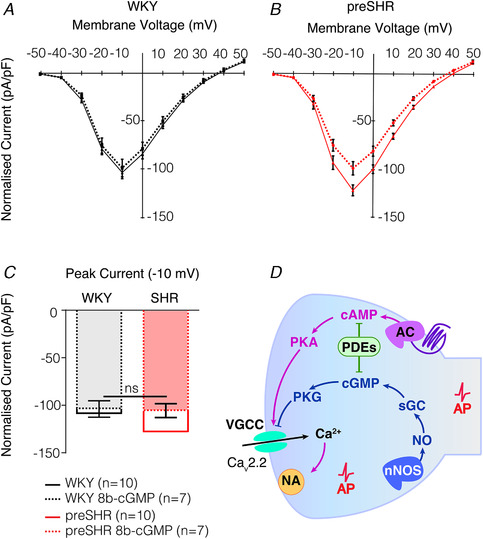
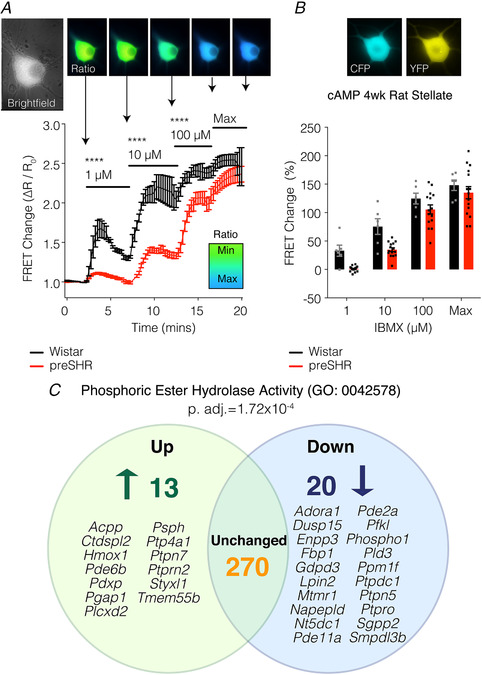
References
-
- Adams MA, Bobik A & Korner PI (1989). Differential development of vascular and cardiac hypertrophy in genetic hypertension. Relation to sympathetic function. Hypertension 14, 191–202. - PubMed
-
- Apparsundaram S & Eikenburg DC (1995). Role of prejunctional beta adrenoceptors in rat cardiac sympathetic neurotransmission. J Pharmacol Exp Ther 272, 519–526. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical