

Purification and Characterization of JZTx-14, a Potent Antagonist of Mammalian and Prokaryotic Voltage-Gated Sodium Channels
- PMID: 30308978
- PMCID: PMC6215091
- DOI: 10.3390/toxins10100408
Purification and Characterization of JZTx-14, a Potent Antagonist of Mammalian and Prokaryotic Voltage-Gated Sodium Channels
Abstract
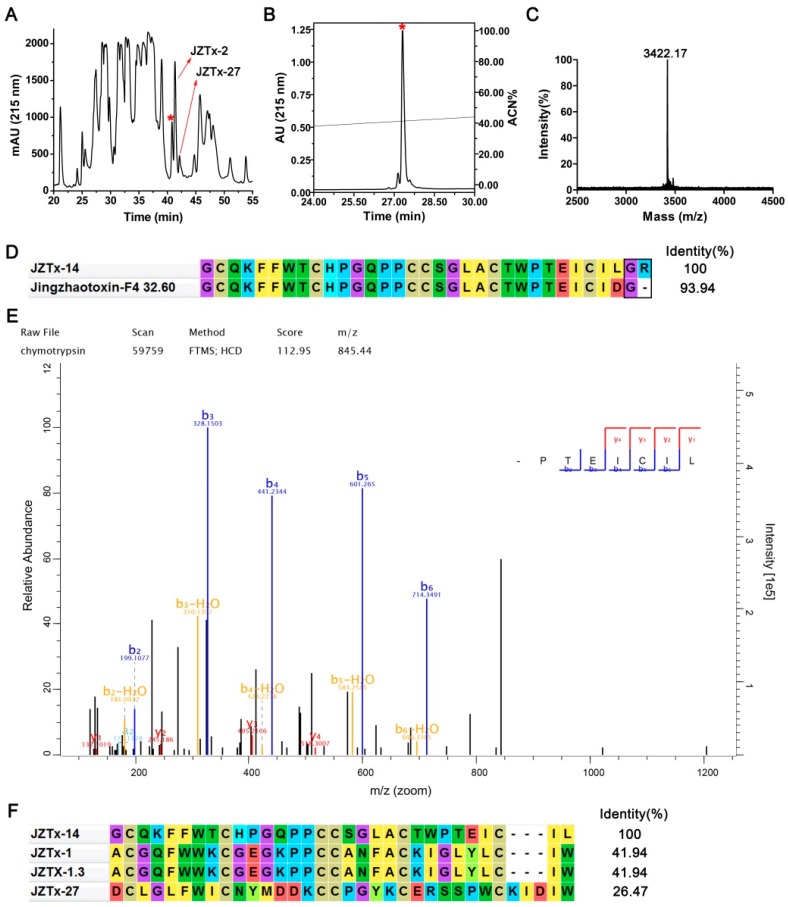
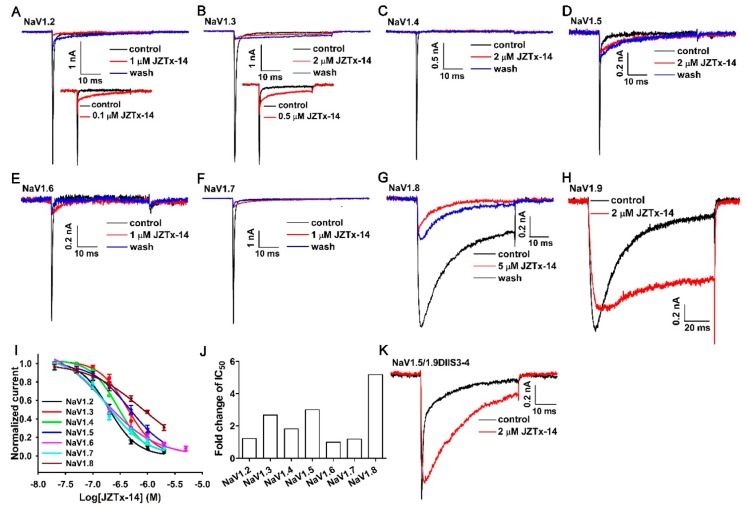
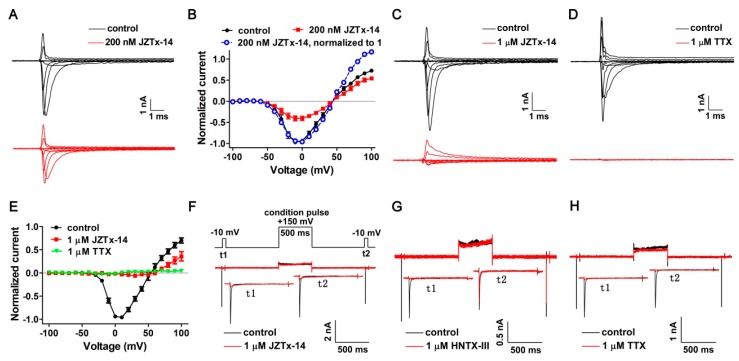
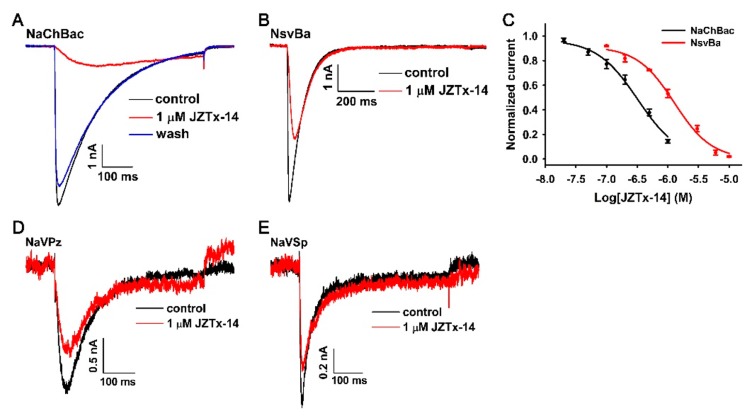
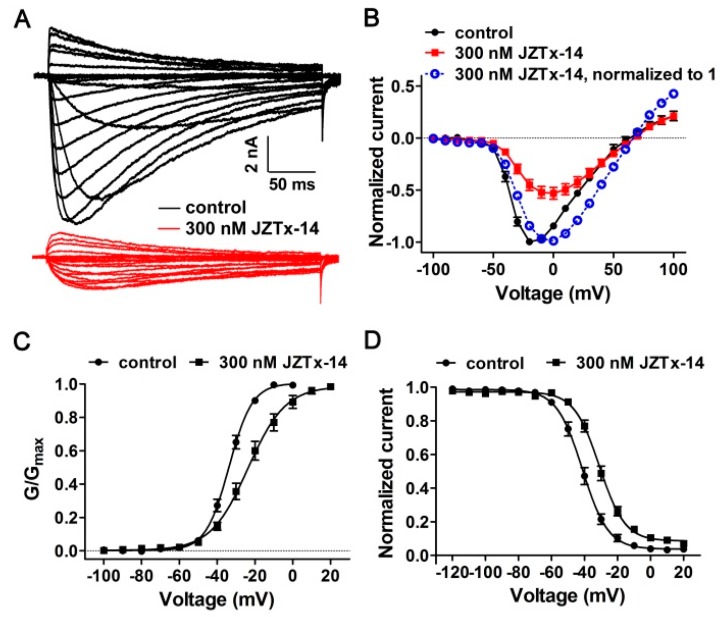
Exploring the interaction of ligands with voltage-gated sodium channels (NaVs) has advanced our understanding of their pharmacology. Herein, we report the purification and characterization of a novel non-selective mammalian and bacterial NaVs toxin, JZTx-14, from the venom of the spider Chilobrachys jingzhao. This toxin potently inhibited the peak currents of mammalian NaV1.2⁻1.8 channels and the bacterial NaChBac channel with low IC50 values (<1 µM), and it mainly inhibited the fast inactivation of the NaV1.9 channel. Analysis of NaV1.5/NaV1.9 chimeric channel showed that the NaV1.5 domain II S3⁻4 loop is involved in toxin association. Kinetics data obtained from studying toxin⁻NaV1.2 channel interaction showed that JZTx-14 was a gating modifier that possibly trapped the channel in resting state; however, it differed from site 4 toxin HNTx-III by irreversibly blocking NaV currents and showing state-independent binding with the channel. JZTx-14 might stably bind to a conserved toxin pocket deep within the NaV1.2⁻1.8 domain II voltage sensor regardless of channel conformation change, and its effect on NaVs requires the toxin to trap the S3⁻4 loop in its resting state. For the NaChBac channel, JZTx-14 positively shifted its conductance-voltage (G⁻V) and steady-state inactivation relationships. An alanine scan analysis of the NaChBac S3⁻4 loop revealed that the 108th phenylalanine (F108) was the key residue determining the JZTx-14⁻NaChBac interaction. In summary, this study provided JZTx-14 with potent but promiscuous inhibitory activity on both the ancestor bacterial NaVs and the highly evolved descendant mammalian NaVs, and it is a useful probe to understand the pharmacology of NaVs.
Keywords: NaChBac; mammalian NaVs; peptide toxin; pharmacology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
