

SRF'ing and SAP'ing - the role of MRTF proteins in cell migration
- PMID: 30309957
- PMCID: PMC6919568
- DOI: 10.1242/jcs.218222
SRF'ing and SAP'ing - the role of MRTF proteins in cell migration
Abstract
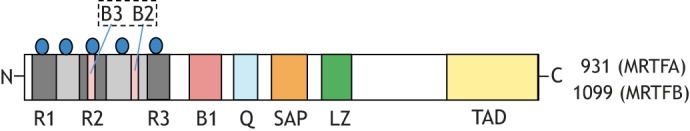
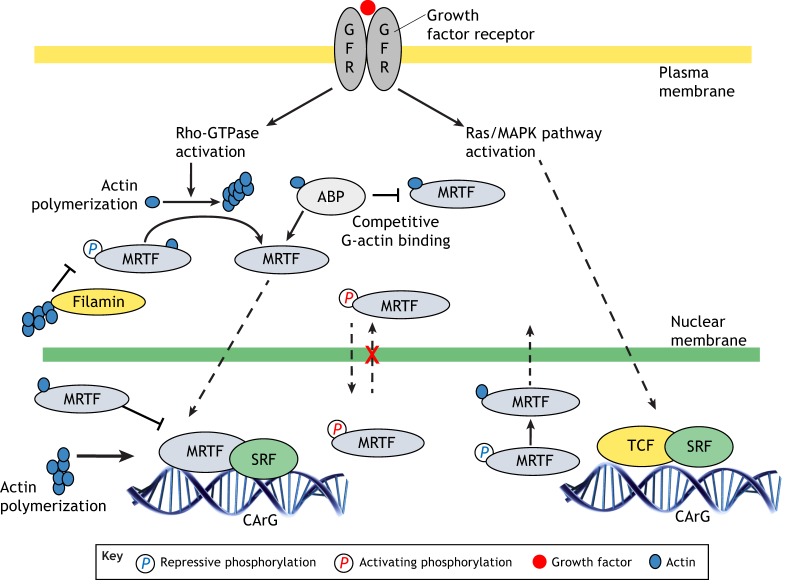
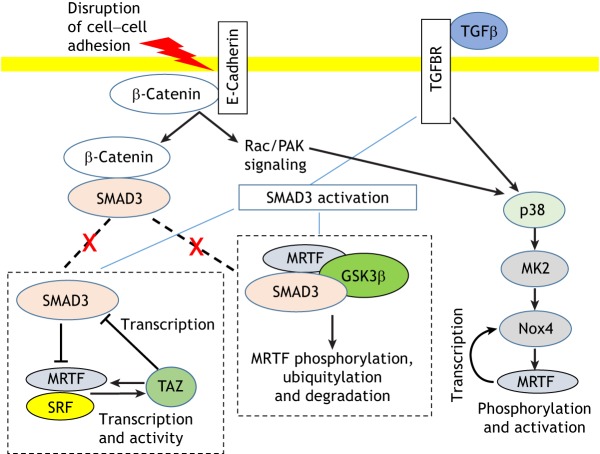
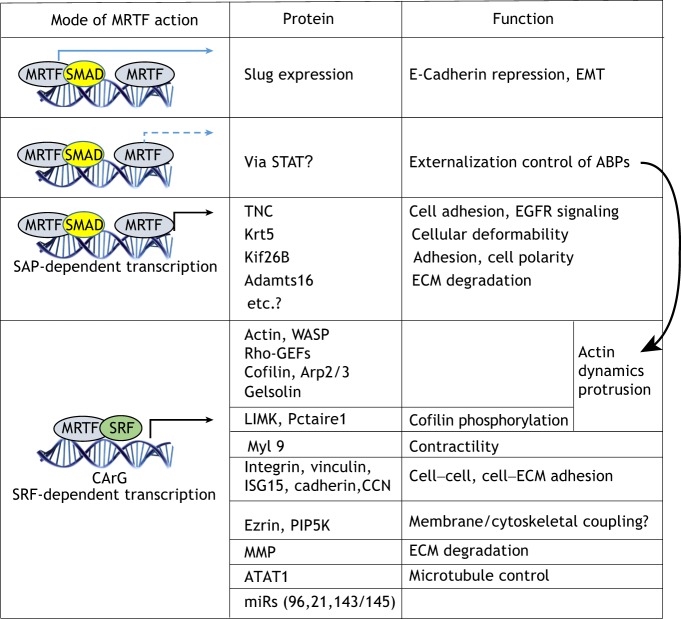
Actin-based cell migration is a fundamental cellular activity that plays a crucial role in a wide range of physiological and pathological processes. An essential feature of the remodeling of actin cytoskeleton during cell motility is the de novo synthesis of factors involved in the regulation of the actin cytoskeleton and cell adhesion in response to growth-factor signaling, and this aspect of cell migration is critically regulated by serum-response factor (SRF)-mediated gene transcription. Myocardin-related transcription factors (MRTFs) are key coactivators of SRF that link actin dynamics to SRF-mediated gene transcription. In this Review, we provide a comprehensive overview of the role of MRTF in both normal and cancer cell migration by discussing its canonical SRF-dependent as well as its recently emerged SRF-independent functions, exerted through its SAP domain, in the context of cell migration. We conclude by highlighting outstanding questions for future research in this field.
Keywords: Actin; Cancer; Cell migration; MRTF; SAP; SRF.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
LMO7 mediates cell-specific activation of the Rho-myocardin-related transcription factor-serum response factor pathway and plays an important role in breast cancer cell migration.Mol Cell Biol. 2011 Aug;31(16):3223-40. doi: 10.1128/MCB.01365-10. Epub 2011 Jun 13. Mol Cell Biol. 2011. PMID: 21670154 Free PMC article.
-
Myocardin-related transcription factor A (MRTF-A) activity-dependent cell adhesion is correlated to focal adhesion kinase (FAK) activity.Oncotarget. 2016 Nov 1;7(44):72113-72130. doi: 10.18632/oncotarget.12350. Oncotarget. 2016. PMID: 27708220 Free PMC article.
-
Muscle-specific signaling mechanism that links actin dynamics to serum response factor.Mol Cell Biol. 2005 Apr;25(8):3173-81. doi: 10.1128/MCB.25.8.3173-3181.2005. Mol Cell Biol. 2005. PMID: 15798203 Free PMC article.
-
Linking actin dynamics and gene transcription to drive cellular motile functions.Nat Rev Mol Cell Biol. 2010 May;11(5):353-65. doi: 10.1038/nrm2890. Nat Rev Mol Cell Biol. 2010. PMID: 20414257 Free PMC article. Review.
-
Actin-mediated gene expression in neurons: the MRTF-SRF connection.Biol Chem. 2010 Jun;391(6):591-7. doi: 10.1515/BC.2010.061. Biol Chem. 2010. PMID: 20370316 Review.
Cited by
-
Histamine-induced biphasic activation of RhoA allows for persistent RhoA signaling.PLoS Biol. 2020 Sep 3;18(9):e3000866. doi: 10.1371/journal.pbio.3000866. eCollection 2020 Sep. PLoS Biol. 2020. PMID: 32881857 Free PMC article.
-
Activation of an actin signaling pathway in pre-malignant mammary epithelial cells by P-cadherin is essential for transformation.Dis Model Mech. 2023 Feb 1;16(2):dmm049652. doi: 10.1242/dmm.049652. Epub 2023 Feb 21. Dis Model Mech. 2023. PMID: 36808468 Free PMC article.
-
The Genomic Response to TGF-β1 Dictates Failed Repair and Progression of Fibrotic Disease in the Obstructed Kidney.Front Cell Dev Biol. 2021 Jul 2;9:678524. doi: 10.3389/fcell.2021.678524. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34277620 Free PMC article. Review.
-
The role of profilin-1 in cardiovascular diseases.J Cell Sci. 2021 May 1;134(9):jcs249060. doi: 10.1242/jcs.249060. Epub 2021 May 7. J Cell Sci. 2021. PMID: 33961053 Free PMC article. Review.
-
MRTF may be the missing link in a multiscale mechanobiology approach toward macrophage dysfunction in space.Front Cell Dev Biol. 2022 Sep 12;10:997365. doi: 10.3389/fcell.2022.997365. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36172272 Free PMC article.
References
-
- Alberti S., Krause S. M., Kretz O., Philippar U., Lemberger T., Casanova E., Wiebel F. F., Schwarz H., Frotscher M., Schutz G. et al. (2005). Neuronal migration in the murine rostral migratory stream requires serum response factor. Proc. Natl. Acad. Sci. USA 102, 6148-6153. 10.1073/pnas.0501191102 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
