

Enzyme Fusion Removes Competition for Geranylgeranyl Diphosphate in Carotenogenesis
- PMID: 30309967
- PMCID: PMC6393812
- DOI: 10.1104/pp.18.01026
Enzyme Fusion Removes Competition for Geranylgeranyl Diphosphate in Carotenogenesis
Abstract
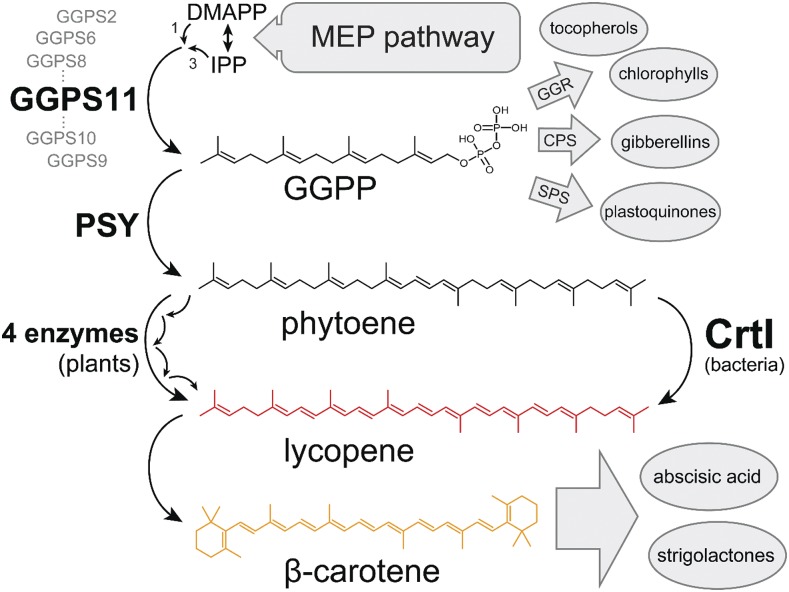
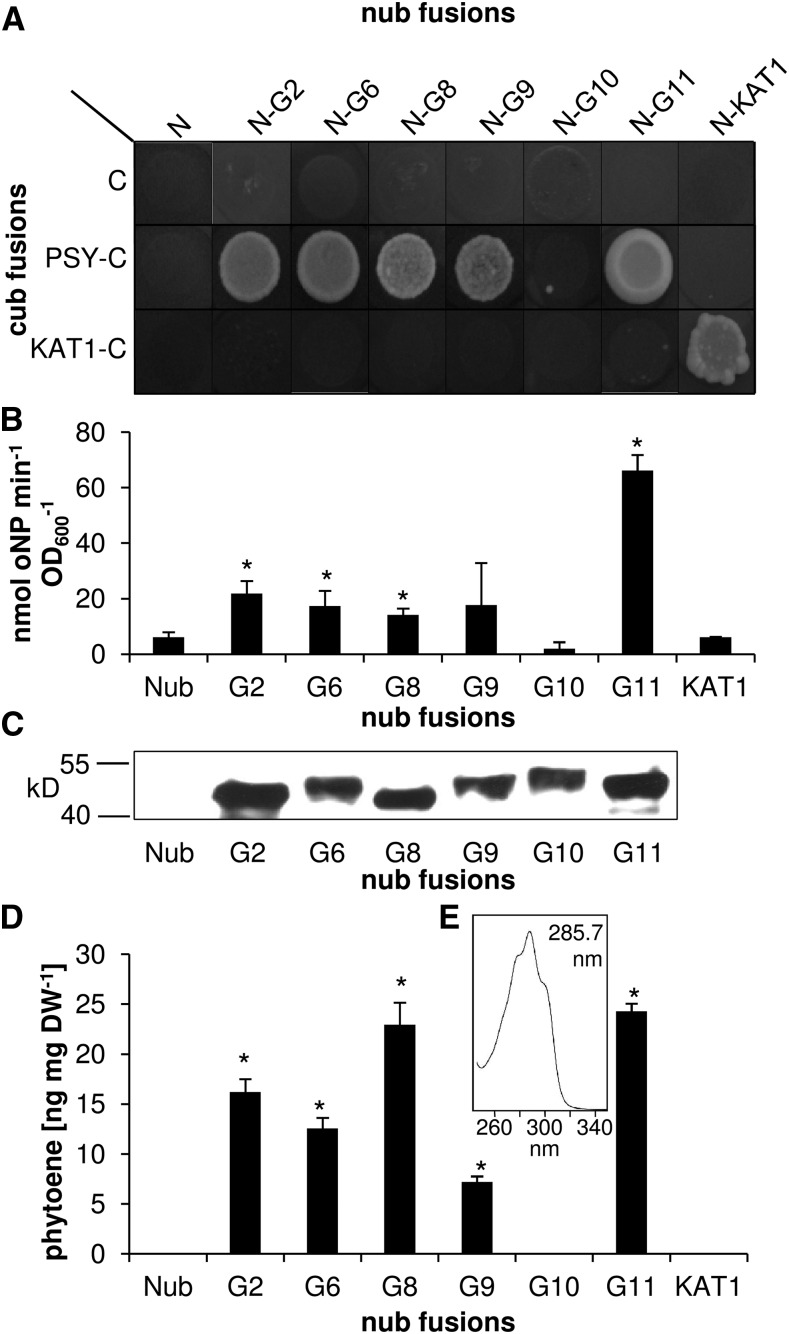
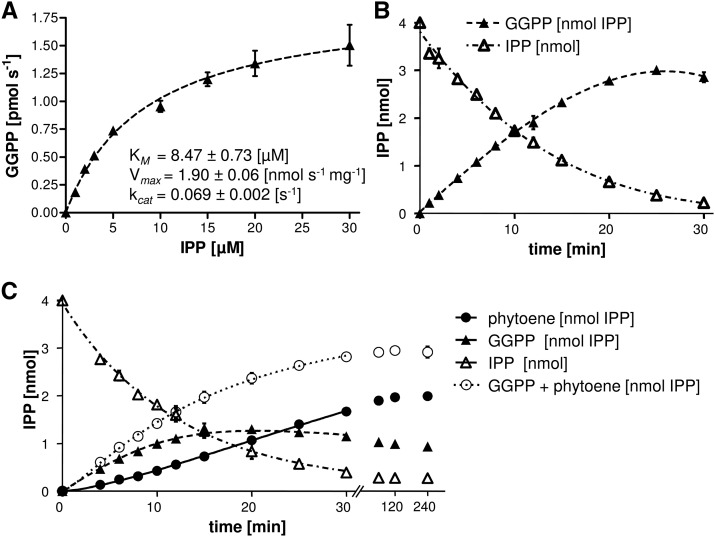
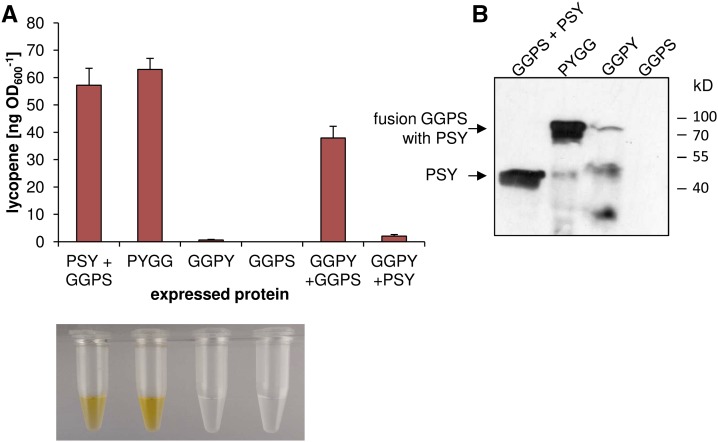
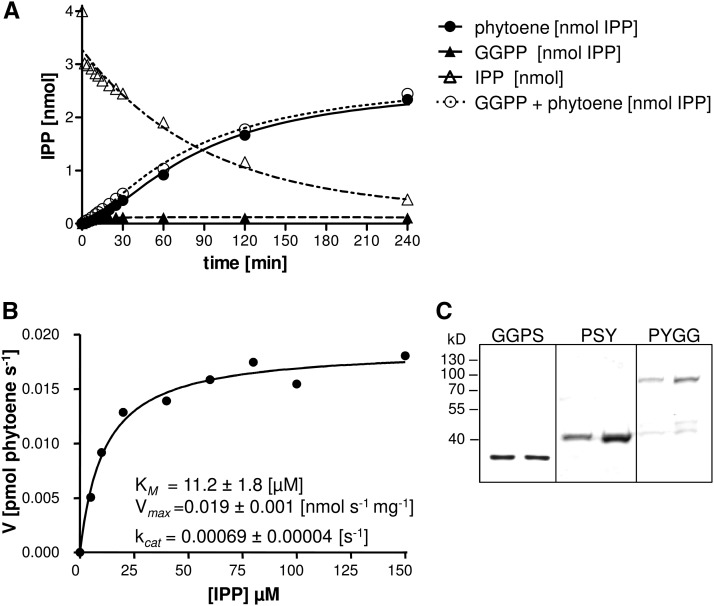
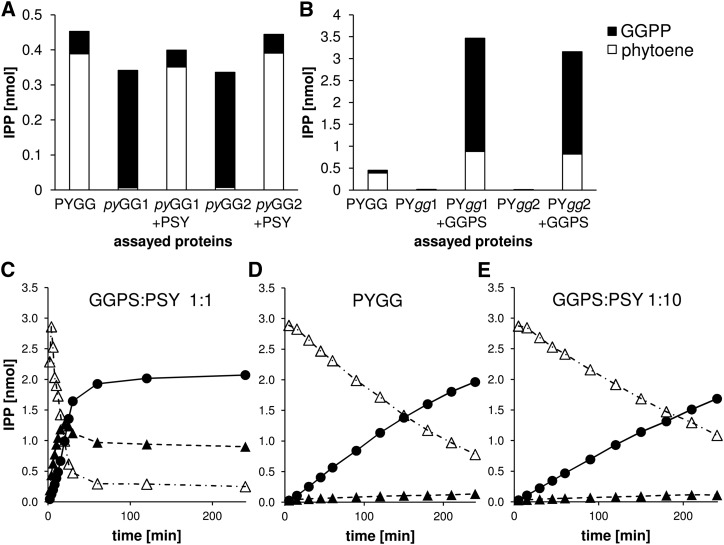
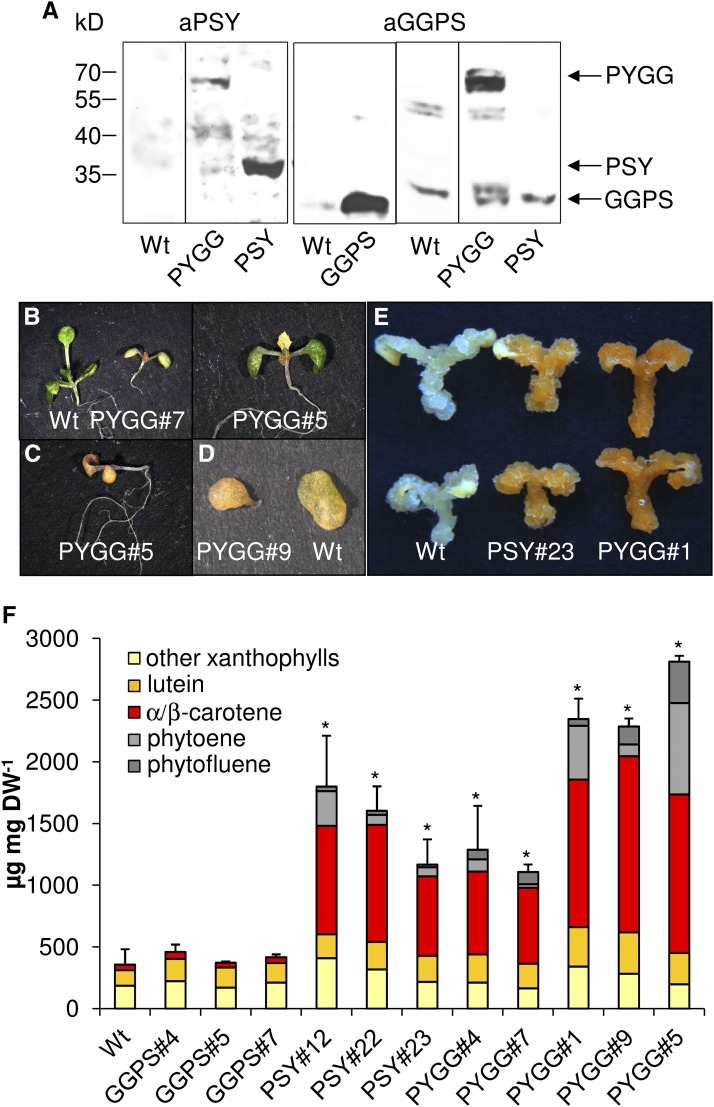
Geranylgeranyl diphosphate (GGPP), a prenyl diphosphate synthesized by GGPP synthase (GGPS), represents a metabolic hub for the synthesis of key isoprenoids, such as chlorophylls, tocopherols, phylloquinone, gibberellins, and carotenoids. Protein-protein interactions and the amphipathic nature of GGPP suggest metabolite channeling and/or competition for GGPP among enzymes that function in independent branches of the isoprenoid pathway. To investigate substrate conversion efficiency between the plastid-localized GGPS isoform GGPS11 and phytoene synthase (PSY), the first enzyme of the carotenoid pathway, we used recombinant enzymes and determined their in vitro properties. Efficient phytoene biosynthesis via PSY strictly depended on simultaneous GGPP supply via GGPS11. In contrast, PSY could not access freely diffusible GGPP or time-displaced GGPP supply via GGPS11, presumably due to liposomal sequestration. To optimize phytoene biosynthesis, we applied a synthetic biology approach and constructed a chimeric GGPS11-PSY metabolon (PYGG). PYGG converted GGPP to phytoene almost quantitatively in vitro and did not show the GGPP leakage typical of the individual enzymes. PYGG expression in Arabidopsis resulted in orange-colored cotyledons, which are not observed if PSY or GGPS11 are overexpressed individually. This suggests insufficient GGPP substrate availability for chlorophyll biosynthesis achieved through GGPP flux redirection to carotenogenesis. Similarly, carotenoid levels in PYGG-expressing callus exceeded that in PSY- or GGPS11-overexpression lines. The PYGG chimeric protein may assist in provitamin A biofortification of edible plant parts. Moreover, other GGPS fusions may be used to redirect metabolic flux into the synthesis of other isoprenoids of nutritional and industrial interest.
© 2019 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Almeida J, Azevedo M da S, Spicher L, Glauser G, vom Dorp K, Guyer L, del Valle Carranza A, Asis R, de Souza AP, Buckeridge M, Demarco D, Bres C, et al. (2016) Down-regulation of tomato PHYTOL KINASE strongly impairs tocopherol biosynthesis and affects prenyllipid metabolism in an organ-specific manner. J Exp Bot 67: 919–934 - PMC - PubMed
-
- Bai C, Twyman RM, Farré G, Sanahuja G, Christou P, Capell T, Zhu C (2011) A golden era—pro-vitamin A enhancement in diverse crops. In Vitro Cell.Dev.Biol.-Plant 47: 1–17 10.1007/s11627-011-9363-6 - DOI
-
- Baillie GS. (2009) Compartmentalized signalling: spatial regulation of cAMP by the action of compartmentalized phosphodiesterases. FEBS J 276: 1790–1799 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
