

Broad phylogenetic analysis of cation/proton antiporters reveals transport determinants
- PMID: 30310075
- PMCID: PMC6181914
- DOI: 10.1038/s41467-018-06770-5
Broad phylogenetic analysis of cation/proton antiporters reveals transport determinants
Abstract
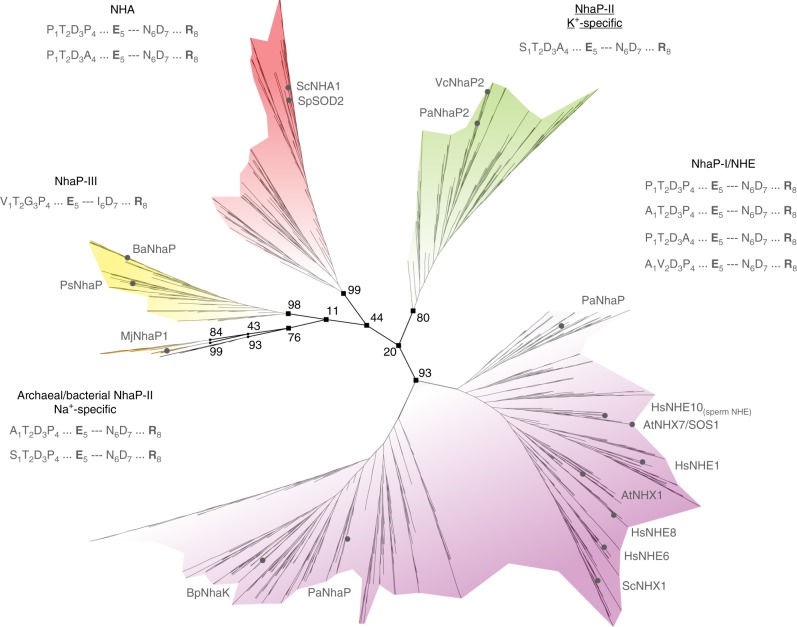
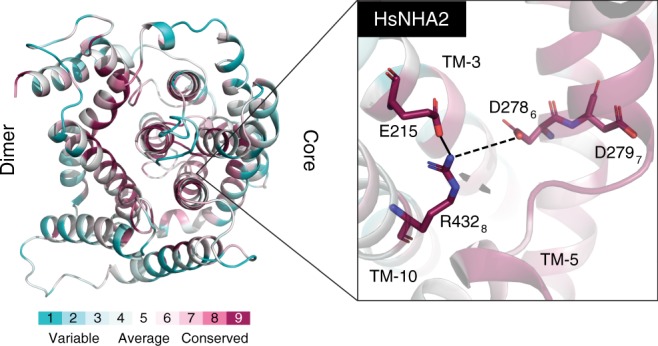
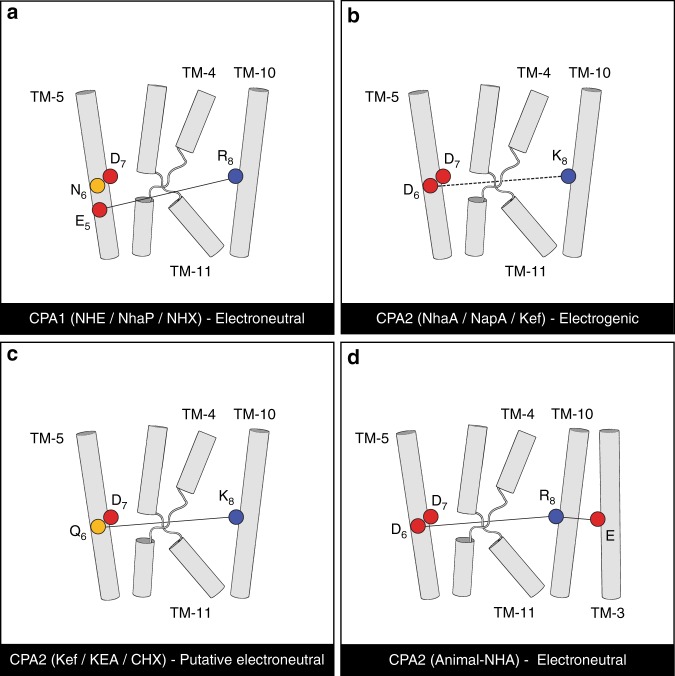
Cation/proton antiporters (CPAs) play a major role in maintaining living cells' homeostasis. CPAs are commonly divided into two main groups, CPA1 and CPA2, and are further characterized by two main phenotypes: ion selectivity and electrogenicity. However, tracing the evolutionary relationships of these transporters is challenging because of the high diversity within CPAs. Here, we conduct comprehensive evolutionary analysis of 6537 representative CPAs, describing the full complexity of their phylogeny, and revealing a sequence motif that appears to determine central phenotypic characteristics. In contrast to previous suggestions, we show that the CPA1/CPA2 division only partially correlates with electrogenicity. Our analysis further indicates two acidic residues in the binding site that carry the protons in electrogenic CPAs, and a polar residue in the unwound transmembrane helix 4 that determines ion selectivity. A rationally designed triple mutant successfully converted the electrogenic CPA, EcNhaA, to be electroneutral.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Yang L, et al. Acute hypertension provokes internalization of proximal tubule NHE3 without inhibition of transport activity. Am. J. Physiol. 2002;282:F730–F740. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
