Ongoing HIV Replication During ART Reconsidered
- PMID: 30310821
- PMCID: PMC6172598
- DOI: 10.1093/ofid/ofx173
Ongoing HIV Replication During ART Reconsidered
Abstract
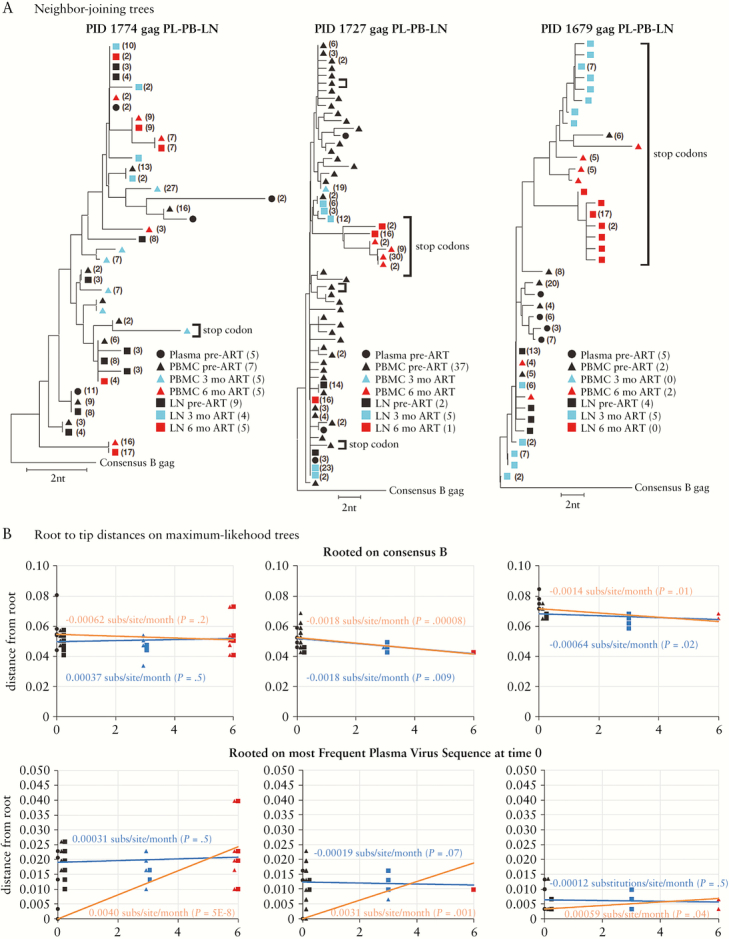
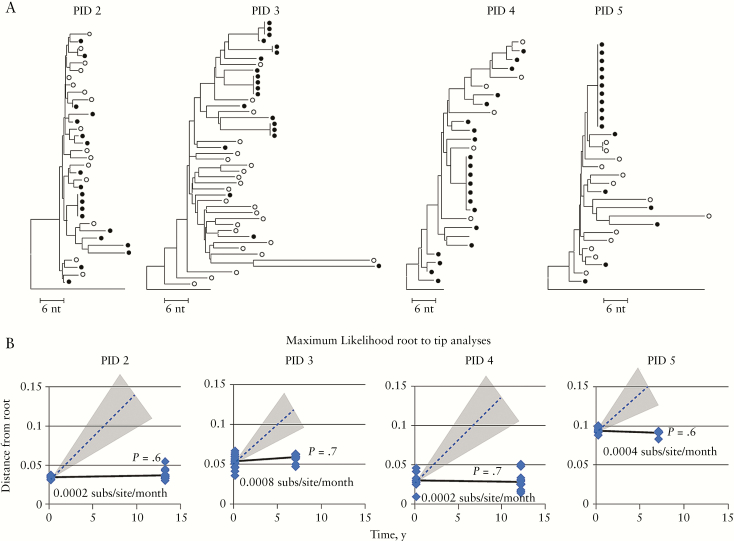
Lorenzo-Redondo et al. recently analyzed HIV RNA sequences in plasma virus and proviral DNA sequences in lymph nodes (LN) and peripheral blood mononuclear cells (PBMC) from samples collected over a 6-month period from 3 individuals following initiation of antiretroviral therapy (ART) and concluded that ongoing HIV replication occurred in LN despite ART and that this replication maintained the HIV reservoir. We analyzed the same sequences and found that the dataset was very limited (median of 5 unique RNA or DNA sequences per sample) after accounting for polymerase chain reaction resampling and hypermutation and that the few remaining DNA sequences after 3 and 6 months on ART were not more diverse or divergent from those in pre-ART in any of the individuals studied. These findings, and others, lead us to conclude that the claims of ongoing replication on ART made by Lorenzo-Redondo et al. are not justified from the dataset analyzed in their publication.
Keywords: HIV evolution; HIV reservoir; clonal expansion; lymph nodes; ongoing replication on ART; single-genome sequencing.
Figures