

High N-glycan multiplicity is critical for neuronal adhesion and sensitizes the developing cerebellum to N-glycosylation defect
- PMID: 30311906
- PMCID: PMC6185108
- DOI: 10.7554/eLife.38309
High N-glycan multiplicity is critical for neuronal adhesion and sensitizes the developing cerebellum to N-glycosylation defect
Abstract
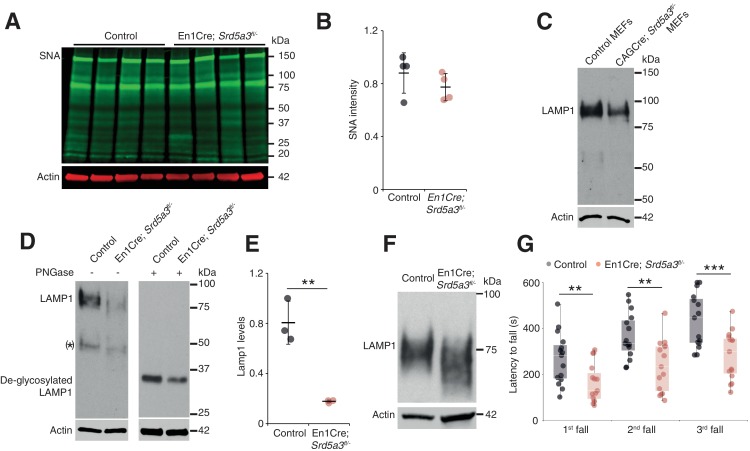
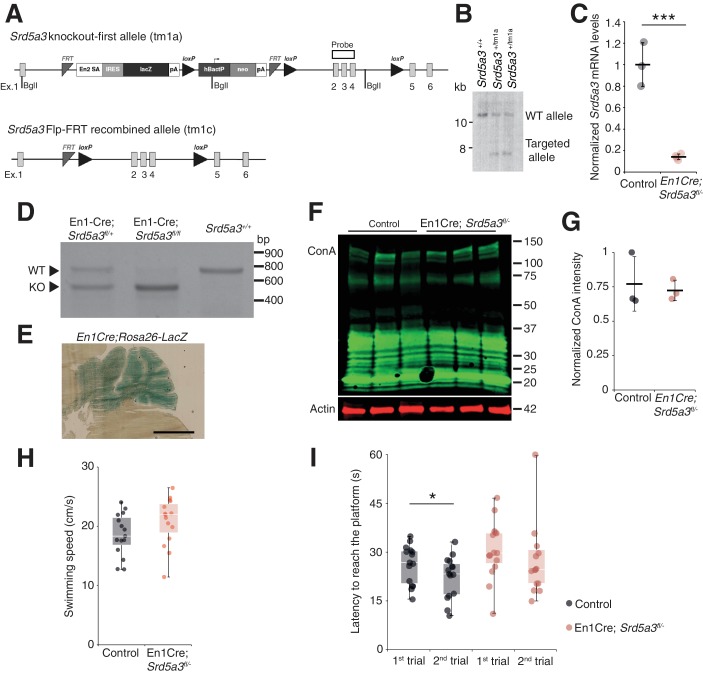
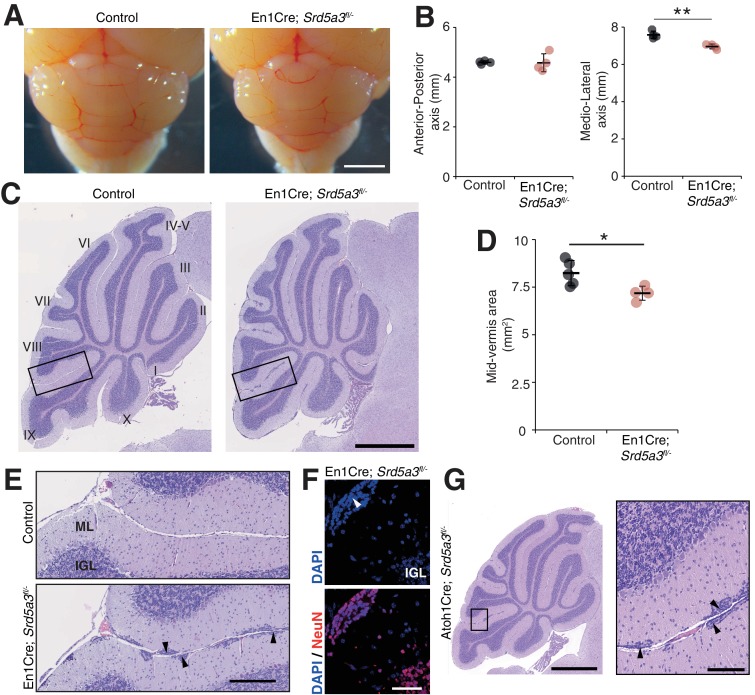
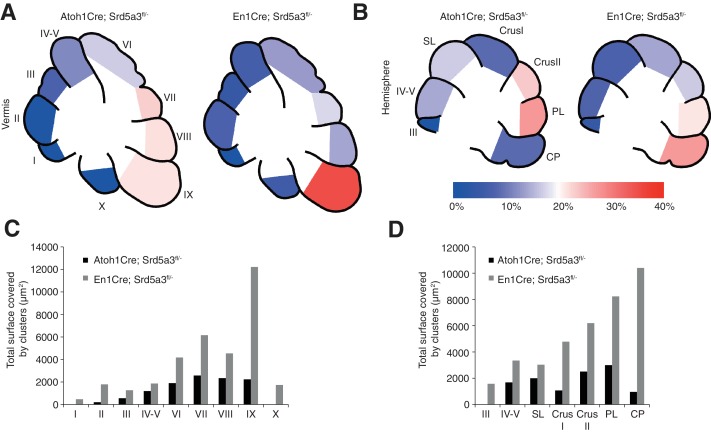
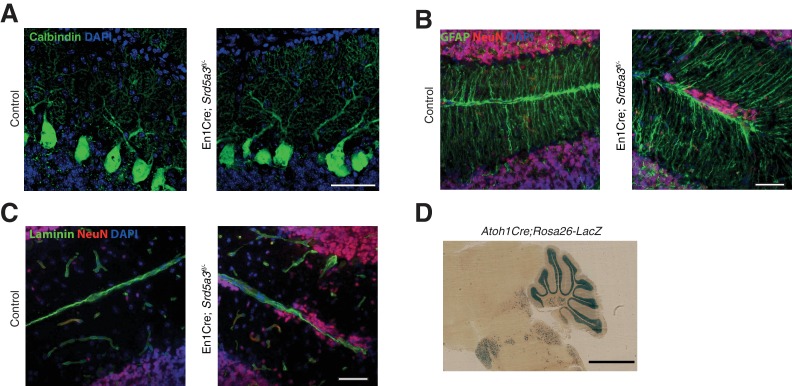
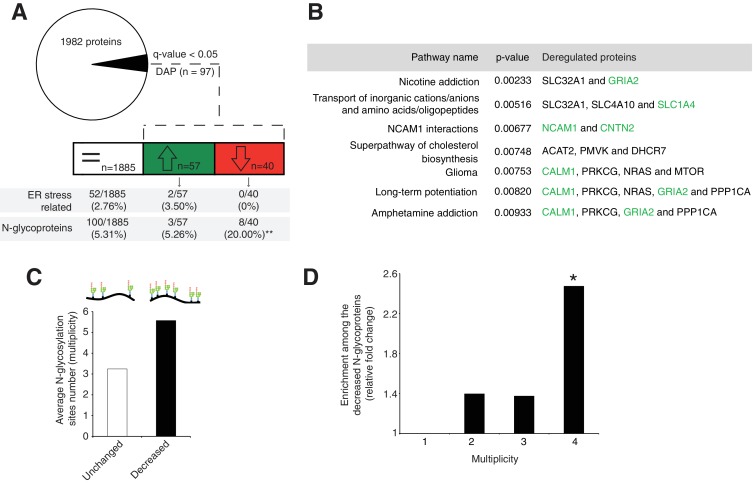
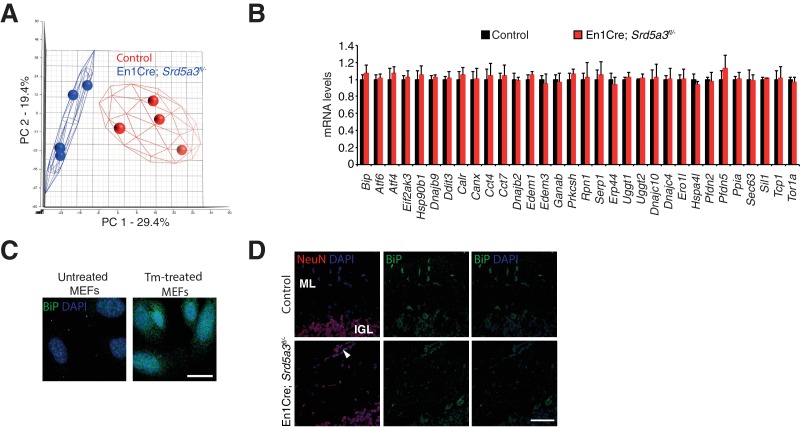
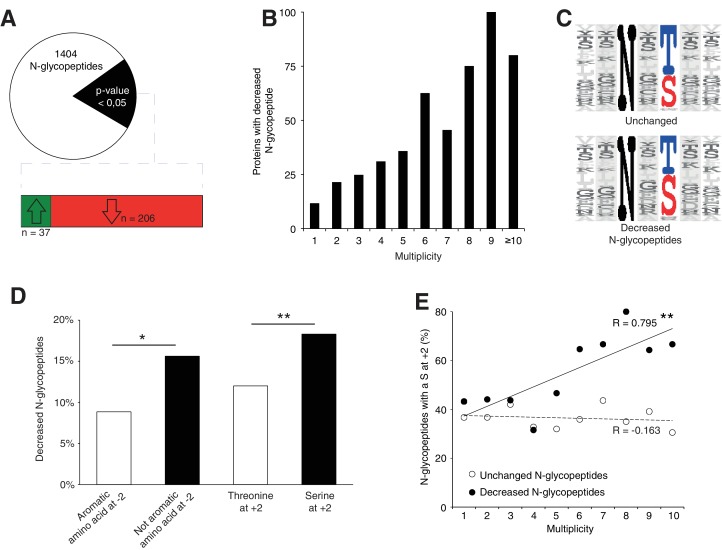
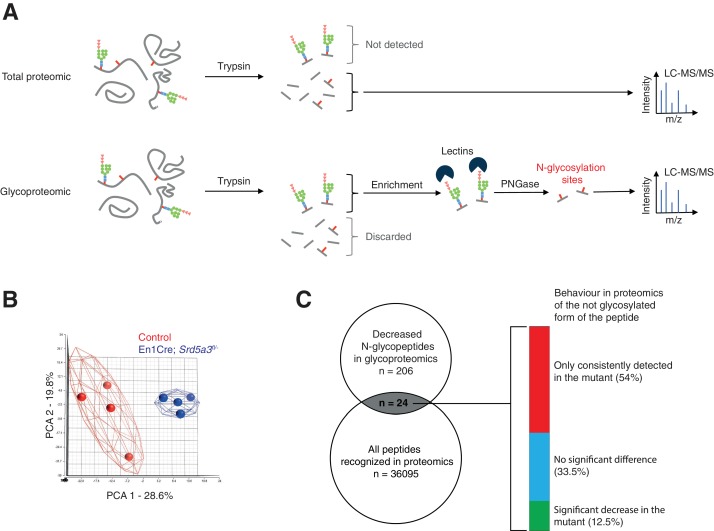
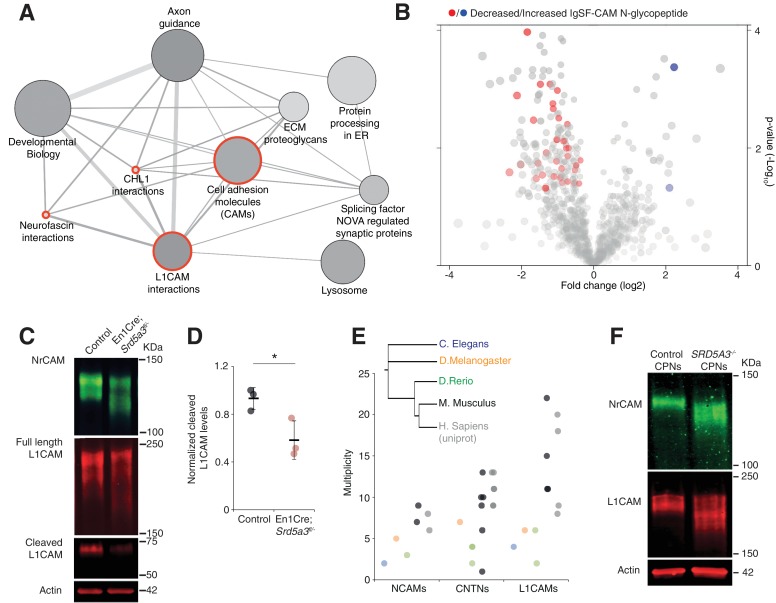
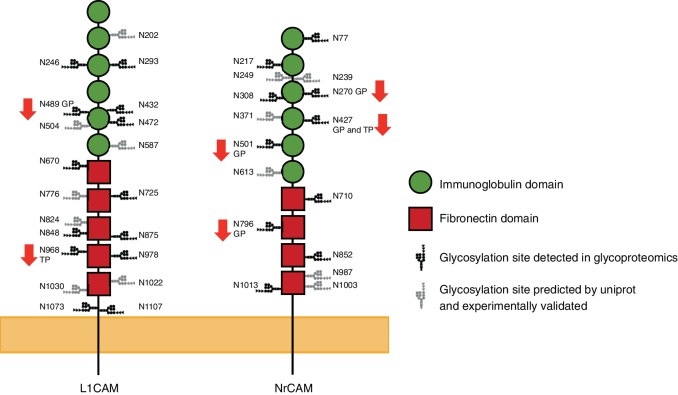
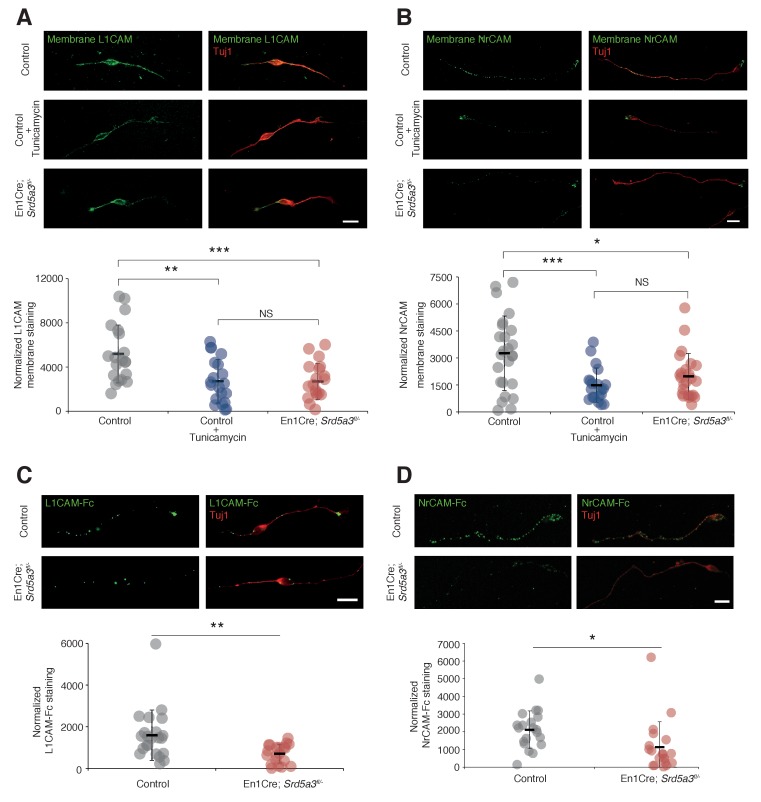
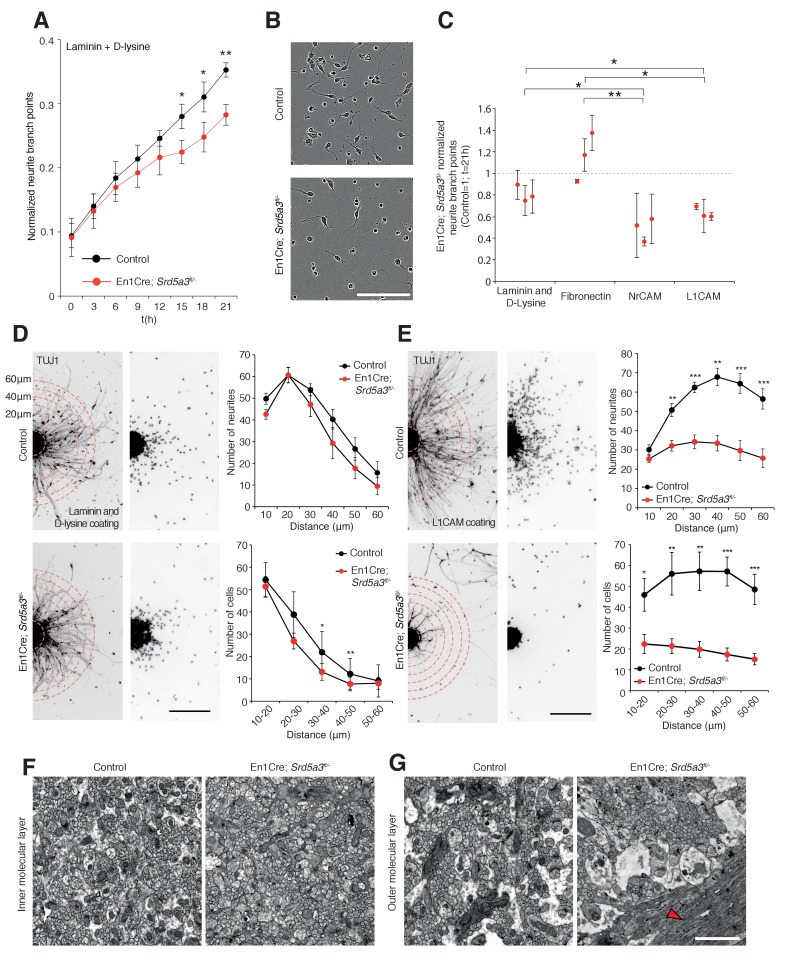
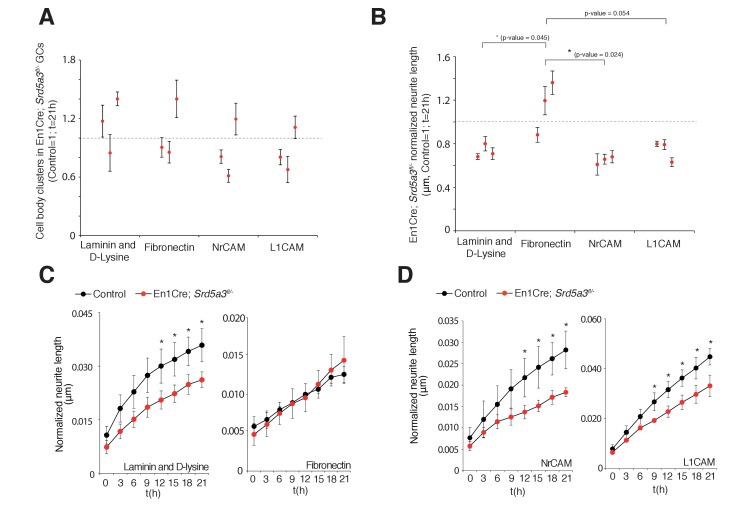
Proper brain development relies highly on protein N-glycosylation to sustain neuronal migration, axon guidance and synaptic physiology. Impairing the N-glycosylation pathway at early steps produces broad neurological symptoms identified in congenital disorders of glycosylation. However, little is known about the molecular mechanisms underlying these defects. We generated a cerebellum specific knockout mouse for Srd5a3, a gene involved in the initiation of N-glycosylation. In addition to motor coordination defects and abnormal granule cell development, Srd5a3 deletion causes mild N-glycosylation impairment without significantly altering ER homeostasis. Using proteomic approaches, we identified that Srd5a3 loss affects a subset of glycoproteins with high N-glycans multiplicity per protein and decreased protein abundance or N-glycosylation level. As IgSF-CAM adhesion proteins are critical for neuron adhesion and highly N-glycosylated, we observed impaired IgSF-CAM-mediated neurite outgrowth and axon guidance in Srd5a3 mutant cerebellum. Our results link high N-glycan multiplicity to fine-tuned neural cell adhesion during mammalian brain development.
Keywords: N-glycosylation; cell adhesion; cell biology; cerebellum; congenital disorders of glycosylation; human; mouse; neuronal migration; neuroscience; proteomics.
© 2018, Medina-Cano et al.
Conflict of interest statement
DM, EU, LN, MN, JL, JB, CT, FF, NL, CF, LC, IG, VC No competing interests declared
Figures
References
-
- Agrawal P, Fontanals-Cirera B, Sokolova E, Jacob S, Vaiana CA, Argibay D, Davalos V, McDermott M, Nayak S, Darvishian F, Castillo M, Ueberheide B, Osman I, Fenyö D, Mahal LK, Hernando E. A systems biology approach identifies FUT8 as a driver of melanoma metastasis. Cancer Cell. 2017;31:804–819. doi: 10.1016/j.ccell.2017.05.007. - DOI - PMC - PubMed
-
- Akizu N, Cantagrel V, Schroth J, Cai N, Vaux K, McCloskey D, Naviaux RK, Van Vleet J, Fenstermaker AG, Silhavy JL, Scheliga JS, Toyama K, Morisaki H, Sonmez FM, Celep F, Oraby A, Zaki MS, Al-Baradie R, Faqeih EA, Saleh MA, Spencer E, Rosti RO, Scott E, Nickerson E, Gabriel S, Morisaki T, Holmes EW, Gleeson JG. AMPD2 regulates GTP synthesis and is mutated in a potentially treatable neurodegenerative brainstem disorder. Cell. 2013;154:505–517. doi: 10.1016/j.cell.2013.07.005. - DOI - PMC - PubMed
-
- Appel F, Holm J, Conscience JF, von Bohlen und Halbach F, Faissner A, James P, Schachner M. Identification of the border between fibronectin type III homologous repeats 2 and 3 of the neural cell adhesion molecule L1 as a neurite outgrowth promoting and signal transducing domain. Journal of Neurobiology. 1995;28:297–312. doi: 10.1002/neu.480280304. - DOI - PubMed
-
- Bizzoca A, Virgintino D, Lorusso L, Buttiglione M, Yoshida L, Polizzi A, Tattoli M, Cagiano R, Rossi F, Kozlov S, Furley A, Gennarini G. Transgenic mice expressing F3/contactin from the TAG-1 promoter exhibit developmentally regulated changes in the differentiation of cerebellar neurons. Development. 2003;130:29–43. doi: 10.1242/dev.00183. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BioSPC program/Université Paris Descartes/International
- ERA-NET 643578-EURO-CDG2/European Commission/International
- ERA-NET E-Rare-3/European Commission/International
- FRM-DEQ20160334938/Fondation pour la Recherche Médicale/International
- ANR-16-CE12- 0005-01/Agence Nationale de la Recherche/International
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
