

Methylation of protein aspartates and deamidated asparagines as a function of blood bank storage and oxidative stress in human red blood cells
- PMID: 30312994
- PMCID: PMC6357231
- DOI: 10.1111/trf.14936
Methylation of protein aspartates and deamidated asparagines as a function of blood bank storage and oxidative stress in human red blood cells
Abstract
Background: Being devoid of de novo protein synthesis capacity, red blood cells (RBCs) have evolved to recycle oxidatively damaged proteins via mechanisms that involve methylation of dehydrated and deamidated aspartate and asparagine residues. Here we hypothesize that such mechanisms are relevant to routine storage in the blood bank.
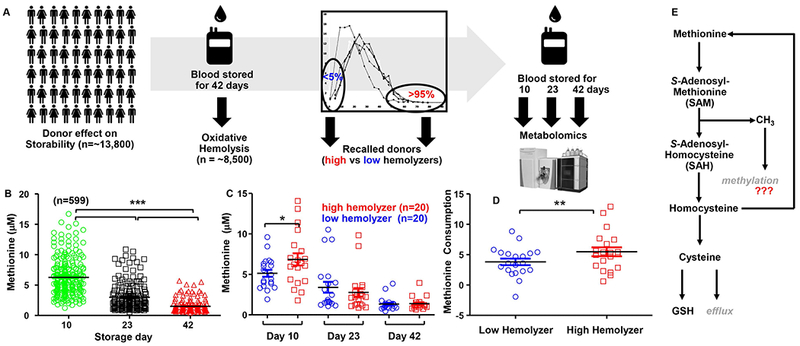
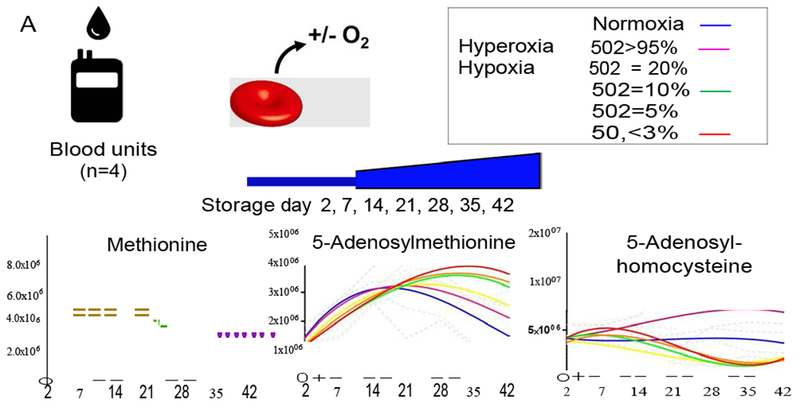
Study design and methods: Within the framework of the REDS-III RBC-Omics (Recipient Epidemiology Donor Evaluation Study III Red Blood Cell-Omics) study, packed RBC units (n = 599) were stored under blood bank conditions for 10, 23, and 42 days and profiled for oxidative hemolysis and time-dependent metabolic dysregulation of the trans-sulfuration pathway.
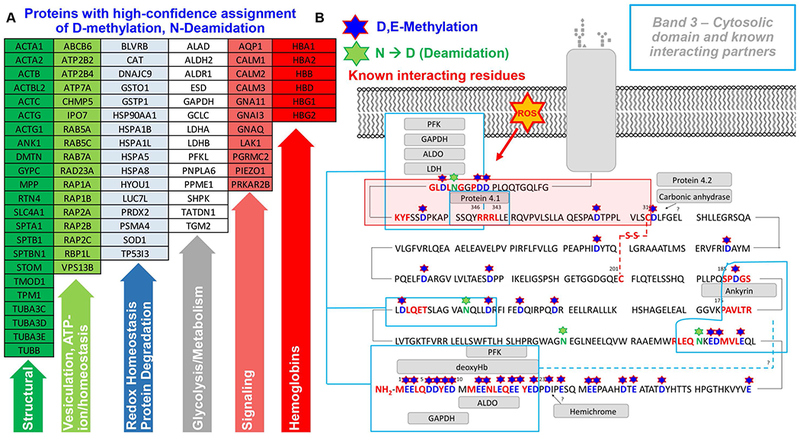
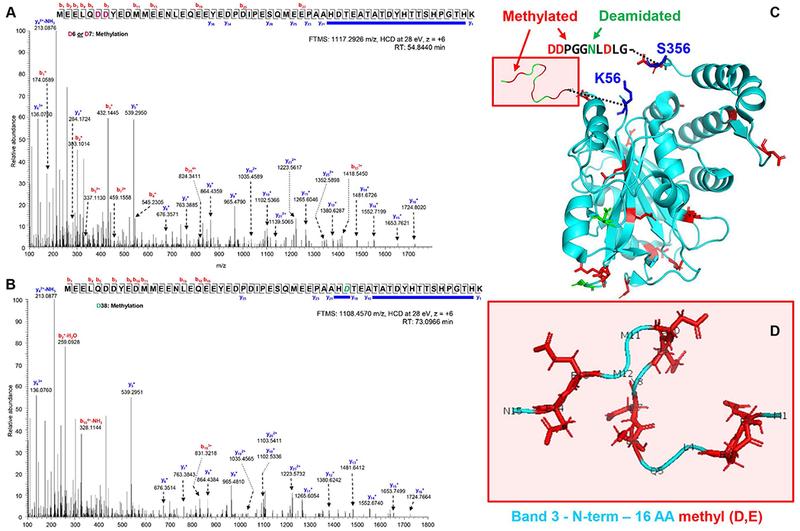
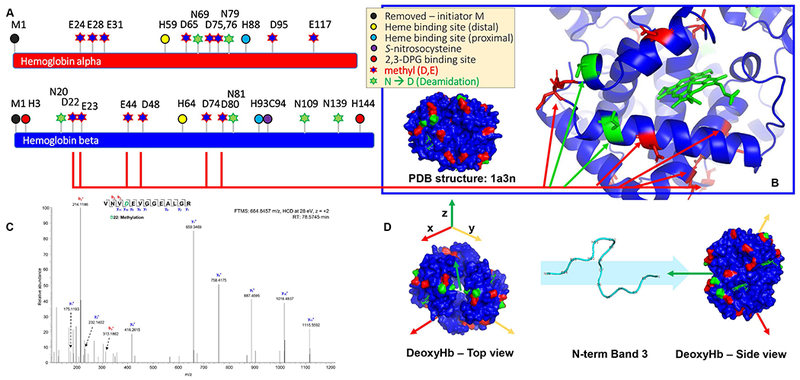
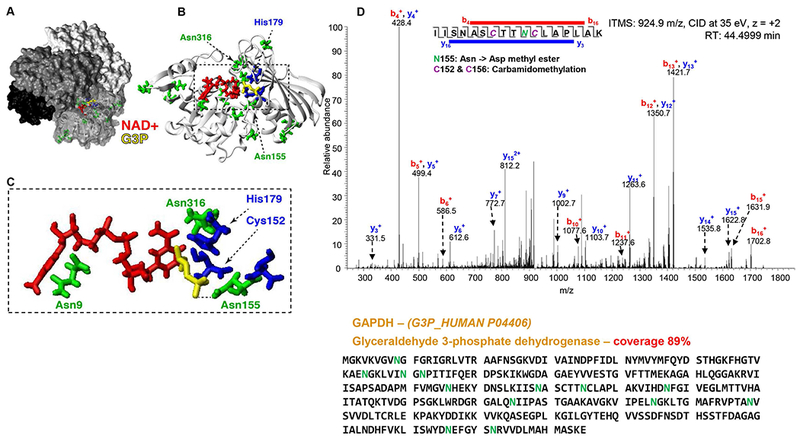
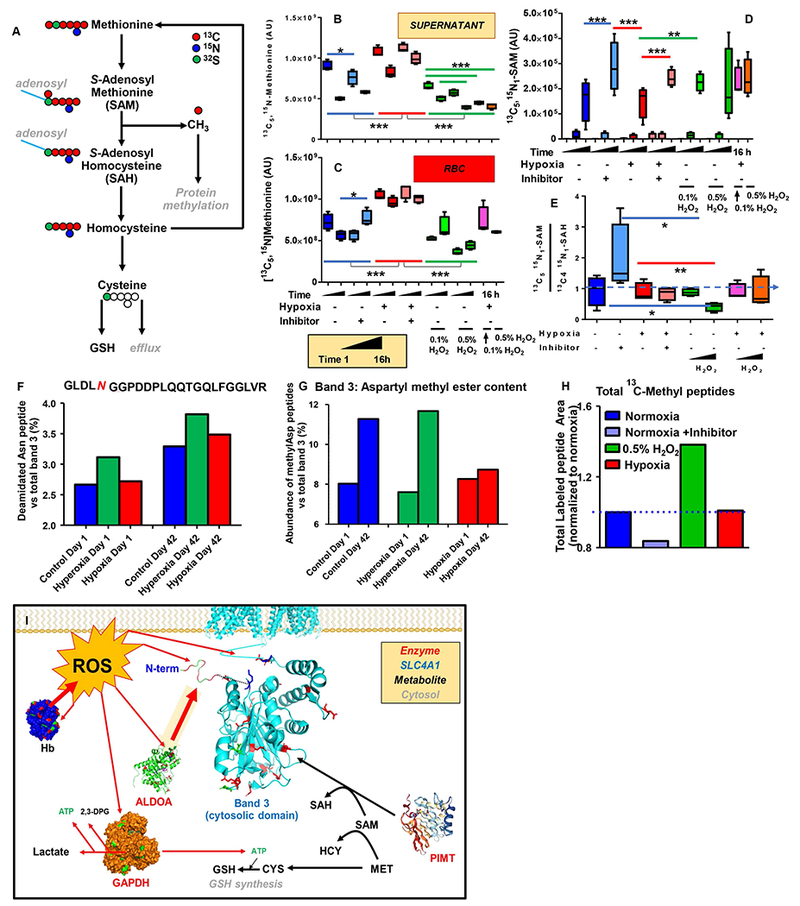
Results: In these units, methionine consumption positively correlated with storage age and oxidative hemolysis. Mechanistic studies show that this phenomenon is favored by oxidative stress or hyperoxic storage (sulfur dioxide >95%), and prevented by hypoxia or methyltransferase inhibition. Through a combination of proteomics approaches and 13 C-methionine tracing, we observed oxidation-induced increases in both Asn deamidation to Asp and formation of methyl-Asp on key structural proteins and enzymes, including Band 3, hemoglobin, ankyrin, 4.1, spectrin beta, aldolase, glyceraldehyde 3-phosphate dehydrogenase, biphosphoglycerate mutase, lactate dehydrogenase and catalase. Methylated regions tended to map proximal to the active site (e.g., N316 of glyceraldehyde 3-phosphate dehydrogenase) and/or residues interacting with the N-terminal cytosolic domain of Band 3.
Conclusion: While methylation of basic amino acid residues serves as an epigenetic modification in nucleated cells, protein methylation at carboxylate side chains and deamidated asparagines is a nonepigenetic posttranslational sensor of oxidative stress and refrigerated storage in anucleated human RBCs.
© 2018 AABB.
Conflict of interest statement
Figures
Comment in
-
Translating red cell "omics" into new perspectives in transfusion medicine: mining the gems in the data mountains.Transfusion. 2019 Jan;59(1):2-5. doi: 10.1111/trf.15066. Transfusion. 2019. PMID: 30615814 No abstract available.
References
-
- D’Alessandro A, Zolla L. Proteomic analysis of red blood cells and the potential for the clinic: what have we learned so far? Expert Rev. Proteomics. 2017;14(3):243–252. - PubMed
-
- Greenwalt TJ A short history of transfusion medicine. Transfusion . 2003;37(5):550–563. - PubMed
-
- D’Alessandro A, Kriebardis AG, Rinalducci S, et al. An update on red blood cell storage lesions, as gleaned through biochemistry and omics technologies. Transfusion . 2015;55(1):205–219. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HHSN268201100002I/HL/NHLBI NIH HHS/United States
- HHSN268201100001I/HL/NHLBI NIH HHS/United States
- HHSN268201100004I/HL/NHLBI NIH HHS/United States
- T32 HL007171/HL/NHLBI NIH HHS/United States
- HHSN263201100003I/NH/NIH HHS/United States
- S10 OD021641/OD/NIH HHS/United States
- HHSN268201100005I/HL/NHLBI NIH HHS/United States
- HHSN268201100008I/HL/NHLBI NIH HHS/United States
- HHSN261201100006I/CA/NCI NIH HHS/United States
- HHSN268201100009I/HL/NHLBI NIH HHS/United States
- HHSN268201100001C/WH/WHI NIH HHS/United States
- HHSN268201100007I/HL/NHLBI NIH HHS/United States
- R03 CA219743/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
